ポイント
- 神経接着因子「DSCAM」をコードする遺伝子の欠損マウス作製に成功
- DSCAM欠損は、中枢性呼吸障害や神経突起伸長の障害を引き起こす
- 呼吸制御メカニズムの理解や中枢性呼吸障害の治療法の開発・改良に大きく寄与
要旨
独立行政法人理化学研究所(野依良治理事長)は、ヒト21番染色体上にあって神経接着因子をコード(暗号化)している遺伝子「DSCAM」のノックアウトマウスを作製し、このマウスが、中枢性呼吸障害(脳にある呼吸中枢の異常による障害)を示し、出生後すぐに死亡することを見いだしました。脳科学総合研究センター(田中啓治センター長代行)神経遺伝研究チームの山川和弘チームリーダー、天野賢治リサーチアソシエイト、記憶学習研究チームの荒田晶子研究員(現:兵庫医科大学生理学・生体機能部門准教授)に加え、同センター神経成長機構研究チーム、分子神経形成研究チーム、行動遺伝学技術開発チーム、小川研究ユニット、および米国ユタ大学などによる共同研究の成果です。
DSCAMは、1998年に山川和弘チームリーダーが、ダウン症精神遅滞の発症にかかわる遺伝子の候補として、ヒト21番染色体上に同定した神経接着因子をコードする遺伝子です(1998年、Human Molecular Genetics)。DSCAMタンパク質は、神経系に広く発現し、神経細胞軸索の誘導など神経回路の形成に関与していると考えられています。研究グループは、この遺伝子のノックアウトマウスを作製し、このマウスが中枢性呼吸障害により、出生後24時間以内に死亡することや、このマウスの神経細胞のDSCAM依存的神経突起伸長が異常を示すことを明らかにしました。これらの成果は、DSCAM遺伝子の異常が、乳幼児突然死症候群※1や中枢性呼吸障害などの疾患の原因の一部となる可能性を示唆するとともに、今後、さらなる呼吸中枢の制御メカニズムの理解や、それら疾患の治療法の開発・改良に寄与することが期待されます。
本研究成果は、北米神経科学会誌『Journal of Neuroscience』(ジャーナル・オブ・ニューロサイエンス)(3月4日号)に掲載されます。
背景
DSCAM遺伝子は、山川和弘チームリーダーが米国留学時(UCLAのジュリー・コーレンバーグ研究室、コーレンバーグ教授は今回の論文の共著者で、現在はユタ大学教授)に、ダウン症精神遅滞の発症にかかわる遺伝子の候補としてヒト21番染色体上に同定したもので、神経接着因子「DSCAM」をコードしています(Yamakawa et al., Human Molecular Genetics, 7: 227-237, 1998)。DSCAMタンパク質は、神経系に広く発現し、神経細胞膜表面で軸索伸長方向を誘導するなど、神経回路の形成に関与していると考えられています。1998年の報告後、ほかの研究プループがハエのDscam遺伝子を同定し、この遺伝子が非常に多くの種類の遺伝子転写産物を作ることや、Dscam遺伝子の完全な欠失が胎生致死となる(出生前に死ぬ)ことを発見してきました(Schmucker et al., Cell, 101: 671-684, 2000)。以来、ハエを用いて、Dscamタンパク質が同類の神経細胞軸索同士の接着を防ぐことや、Dscam遺伝子発現の減少や過剰が神経ネットワークの異常をもたらすことなど、数多くの研究成果が発表されてきました。マウスでも、Dscam遺伝子自然欠失マウスにおける網膜神経細胞軸索伸長とモザイク構造の異常(Fuert et al., Nature 451:470-474, 2008)などの報告があり、その中で当該マウスが出生後に急死する記載もありましたが、その理由は分かっていませんでした。
研究手法と成果
研究グループは、相同組み換え法※2によってマウスDscam遺伝子をノックアウトし、ヘテロ接合体※3(+/-)を作製しました。変異を導入したヘテロ接合体(+/-)マウス同士を掛け合わせたところ、野生型(+/+)、ヘテロ接合体(+/-)、ホモ接合体※3(-/-)のマウスがほぼ通常の割合(1:2:1)で生まれました。
これらのマウスの脳の遺伝子転写産物(mRNA)を、ノーザンブロット解析※4で解析したところ、Dscam遺伝子転写産物はヘテロ接合体(+/-)マウスでは半減し、ホモ接合体(-/-)マウスでは消失していることを確認しました(図1)。
ヘテロ接合体(+/-)、ホモ接合体(-/-)マウスは、ともに外見上はほぼ正常に生まれてきます。しかし、ホモ接合体(-/-)マウスは、出生後ほとんどすべてが、24時間以内に苦しくもがくようなしぐさをして急死します。プレシスモグラフィー※5により呼吸を測定したところ、野生型(+/+)に比べて、ヘテロ接合体(+/-)では呼吸リズム、呼吸深度の乱れが見られ、ホモ接合体(-/-)ではその乱れがさらに激しくなり、無呼吸も見られることが分かりました(図2)。
この呼吸の乱れが、中枢性のものか肺などの末梢性のものかを判別するため、研究グループは、新生マウス摘出脳幹-脊髄標本※6を用いて、呼吸性神経回路網を光学的測定法により解析しました。延髄に位置し、呼吸リズムを生み出すとされる2つの呼吸中枢(傍顔面神経核呼吸性ニューロン群:pFRGおよびプレベッチンガー複合体:Pre-Böt)の活動を、電位依存性の色素を用いて経時的に解析したところ、野生型(+/+)に比べてヘテロ接合体(+/-)では、吸息先行型ニューロンが多いとされるpFRGの光学的シグナルが減少し、ホモ接合体(-/-)では光学的シグナルが消失することが分かりました。その一方、吸息性ニューロンが多いとされるPre-Bötの活動は、ヘテロ接合体(+/-)およびホモ接合体(-/-)でも比較的光学的シグナルが保たれていました(図3)。
また、野生型(+/+)およびホモ接合体(-/-)マウスの後根神経節※7を摘出し、DSCAMタンパク質を塗布した培養皿に静置して神経突起伸長の長さを測定したところ、ホモ接合体(-/-)マウス後根神経節の突起伸長が、野生型(+/+)マウスのそれに比べて低下していることが分かりました(図4)。これは、DSCAMがDSCAM-DSCAM接着依存的な神経突起伸長活性を担い、ホモ接合体(-/-)マウスではその活性が失われていることを示しています。
これらの結果は、Dscam遺伝子の欠損が、呼吸中枢の1つである吸息先行型ニューロンネットワークを撹乱させ、そのネットワーク同調の異常をもたらすことによって呼吸リズムに異常を来たし、出生後まもなく急死を引き起こすことを示唆します。さらにこれらの成果は、いまだにその細部が明らかでない、呼吸中枢の制御メカニズムの理解につながるとともに、DSCAM遺伝子の異常が、乳幼児突然死症候群や中枢性呼吸障害などの疾患の原因の一部となる可能性をも示しました。
今後の期待
今回の知見と作製したモデルマウスは、今後、乳幼児突然死症候群や中枢性呼吸障害などの疾患の治療法の開発・改良に大きく寄与することが期待されます。また、ダウン症に見られる呼吸異常では、器質的な異常(筋緊張低下など)に加え、中枢性呼吸制御機構の異常も示唆されていることや、DSCAMタンパク質が、呼吸中枢の存在する延髄ばかりでなく、記憶学習の役割を担う大脳、海馬などにも広く発現しています。これらのことから、DSCAMタンパク質がダウン症の呼吸障害や精神遅滞の発症にかかわっている可能性もあり、その役割の解明にも今後役立つものと強く期待されます。
発表者
理化学研究所
脳科学総合研究センター 神経遺伝研究チーム
チームリーダー 山川 和弘
お問い合わせ先
脳科学研究推進部 鈴木 一郎(すずき いちろう)Tel : 048-467-7526 / Fax : 048-462-4914
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.乳幼児突然死症候群
検死解剖、死因調査、加療歴などすべての情報を精査しても原因が特定できない、一見、健康に見える乳児の突然死。生後1週間から1歳までの乳児死亡の原因では最も多く、出生1,000人あたり約2人の割合で起こる。 - 2.相同組み換え法
標的となる遺伝子に変異を導入したDNAを、マウス細胞に導入して相同組み換えを起こさせることで、変異をゲノムDNAに導入する方法。この方法により、マウスES細胞(胚性幹細胞)に変異や欠失を導入し、ノックアウトマウスやノックインマウスなどが作製できる。 - 3.ヘテロ接合体、ホモ接合体
遺伝子は1個体当たり、父親由来と母親由来の2コピー存在する。ヘテロ接合体の場合では、正常遺伝子1コピーと変異遺伝子1コピー、ホモ接合体の場合では、変異遺伝子2コピーを有する。 - 4.ノーザンブロット解析
組織などからの遺伝子転写産物(RNA:リボ核酸)を電気泳動し、膜に移した後、標的転写産物に相補性のある核酸プローブで標識し解析する方法。 - 5.プレシスモグラフィー
マウスなどの動物を筒状の容器に入れ、内部の気圧変化を測定することにより呼吸リズムを解析する方法。 - 6.摘出脳幹-脊髄標本
新生ラットやマウスの脳幹と脊髄を取り出してチェンバーにてリンゲル液でかん流しながら生かす標本。血液かん流がなく、求心性入力のない状態で頚髄の第4前根から呼吸リズムが記録できる。 - 7.後根神経節
脊髄神経知覚枝の根部にある神経節(神経細胞体が集積した構造物)。

図1 野生型マウス、Dscam変異ヘテロ接合体、ホモ接合体の脳遺伝子転写産物のノーザンブロット解析
Dscam 遺伝子転写産物(9.5kb)は、ヘテロ接合体(+/-)では半減し、ホモ接合体(-/-)マウスでは消失していた。
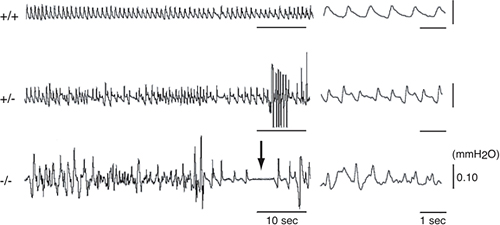
図2 Dscam変異ヘテロ接合体およびホモ接合体でみられた呼吸の異常野生型(+/+); ヘテロ接合体(+/-); ホモ接合体(-/-)
プレシスモグラフィーで測定した呼吸リズム。野生型に比べて、ヘテロ接合体では呼吸の乱れが見られ、ホモ接合体ではさらに乱れが激しくなっている。矢印は無呼吸を示す。
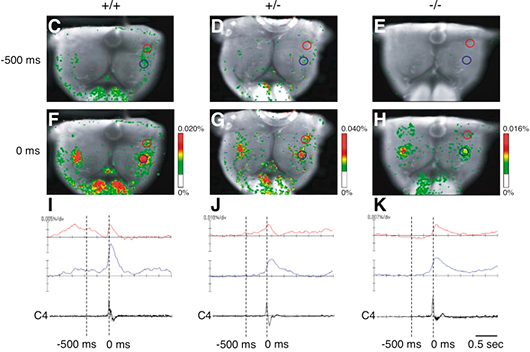
図3 電位依存性色素を用いた新生マウス摘出脳幹―脊髄標本における呼吸中枢の解析
野生型(+/+); ヘテロ接合体(+/-); ホモ接合体(-/-)
赤丸と赤線:pFRGの位置とその発火、
青丸と青線:pre-Bötの位置とその発火
標本の吻側に位置する赤丸印のところは、傍顔面神経核呼吸性ニューロン群(pFRG)で、主に吸息先行型ニューロンが多く見られるため、吸息活動が始まる前から光学シグナルが観察される。標本の青丸印のところは、プレベッチンガー複合体(pre-Böt)と呼ばれ、pFRGよりもやや尾側に位置し、吸息性ニューロンが多く見られるところとなる。それゆえ、光学的測定法により、時間的・空間的に分かれて見えるのである。野生型(C、F、I)に比べて、ヘテロ接合体(D、G、J)およびホモ接合体(E、H、K)では、-500msにおけるpFRGの光学的シグナルの減少、消失が見られる。ヘテロ接合体(D、G、J)およびホモ接合体(E、H、K)においてもpre-Bötの光学的シグナルは見られるが、野生型に比べて弱く、またそのシグナルは大きく損なわれている(図2参照)。(注:野生型において、-500msにおけるpFRGの発火は0msにおけるpre-Bötの活動を一部誘導制御し、pre-Bötの活動は横隔膜を駆動するC4神経活動(第4頚髄前根の神経活動)を直接制御する。)
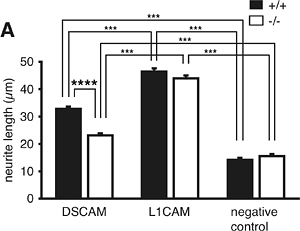
図4 マウス摘出後根神経節を用いたDSCAM-DSCAM接着依存的神経突起伸長の解析
野生型(+/+)およびホモ接合体(-/-)マウスの後根神経節を摘出し、DSCAMタンパク質、もしくはL1CAMタンパク質(別の神経接着因子:比較対象として使用した)を塗布した培養皿に静置して7時間観察し、神経突起伸長の長さを測定した。DSCAMタンパク質を塗布した培養皿において、ホモ接合体(-/-)マウス後根神経節の突起伸長が野生型(+/+)マウスのそれに比べ、有意に低下している。(negative control:神経接着因子の塗布なし。)