ポイント
- ゼブラフィッシュの嗅覚神経回路を緑色蛍光タンパク質「GFP」で可視化
- 嗅覚中枢から、感情を調節する神経核につながる左右非対称な回路を発見
- 右利き、左利きなど動物行動の左右非対称性を理解する新たな知見
要旨
独立行政法人理化学研究所(野依良治理事長)は、モデル動物で知られる熱帯魚ゼブラフィッシュ※1の脳の神経回路を緑色蛍光タンパク質で可視化し、嗅覚系に左右非対称な神経回路が存在することを発見しました。理研脳科学総合研究センター(利根川進センター長)シナプス分子機構研究チームの吉原良浩チームリーダーと宮坂信彦 副チームリーダー、発生遺伝子制御研究チームの岡本仁チームリーダーおよび自然科学機構・岡崎統合バイオサイエンスセンター(永山國昭センター長)による共同研究の成果です。
匂い(におい)の源から発せられた「匂い分子」は、鼻の奥にある感覚神経細胞(嗅細胞)が受け取り、その匂い情報を、神経線維を介して脳の入り口の嗅球※2に伝えます。動物が、匂いを認識して「快・不快」あるいは「好き・嫌い」の感情を表現するためには、匂いの情報をさらに高次の嗅覚中枢へと伝え、処理する必要があります。しかし、嗅球の神経細胞(僧帽細胞※3)が、高次中枢とどのような神経ネットワークを形成しているのかについては、ほとんど分かっていませんでした。
研究チームは、遺伝子工学的手法を用いてゼブラフィッシュの僧帽細胞に緑色蛍光タンパク質GFP(2008年 下村脩博士ら ノーベル化学賞受賞)を発現させ、僧帽細胞の神経線維を可視化し、高次中枢との接続様式を詳細に解析しました。その結果、ある一群の僧帽細胞は、神経線維を左右の終脳※4に対称に投射するとともに、右の手綱核※5(たづなかく)だけに投射することを発見しました。手綱核は、喜び・恐怖などの感情(情動)に深くかかわるとされている脳の部位です。今回の成果により、嗅球と手綱核を直接結ぶ神経接続の存在を初めて明らかにし、匂いによって呼び起こされる情動のメカニズムを解明する糸口を見いだしました。また、発見した神経回路が左右非対称であることは、動物行動の非対称性(例えば、右利き・左利きなど)の理解に大きく貢献するものと期待されます。
研究は、文部科学省特定領域研究「細胞感覚」および同特定領域研究「細胞外環境」の助成金を得て実施し、研究成果は米国の科学雑誌『Journal of Neuroscience』オンライン版(4月15日号)に掲載されます。
背景
嗅覚は、多くの動物で、外界の情報を感知するセンサーとしての機能を持ち、摂食行動、危険回避行動、繁殖行動などの生命活動に重要な役割を果たしています。2004年のノーベル医学生理学賞は、鼻の奥にある感覚神経細胞(嗅細胞)の嗅覚受容体※6遺伝子を発見した米国のリチャード・アクセル(Richard Axel)博士とリンダ・バック(Linda Buck)博士に贈られました。両博士の発見(1991年)が契機となって、匂いの受容メカニズムと鼻から脳への神経配線様式の理解は、その後飛躍的に進みました。
「匂い」の成分であるさまざまな化学物質(匂い分子)は、嗅細胞の嗅覚受容体と結合します。嗅覚受容体遺伝子は、ヒトでは約350種類、マウスでは約1,300種類、魚では約200種類ほど存在していますが、個々の嗅細胞は、その中からたった一種類の嗅覚受容体を選択して発現し、特定の化学構造を持つ匂い分子だけを認識します。匂いの情報は、嗅細胞の神経線維を介して、脳の先端の嗅球と呼ばれる最初の情報処理中枢に伝わります(図1)。同じ受容体を持つ嗅細胞の神経線維は、嗅球表面に並ぶ多くの糸球体※7(匂い情報処理のユニット構造)のうちの特定の糸球体と接続しています。すなわち、多種多様な匂い分子の情報は、糸球体の配置によって規定される「匂い地図」として嗅球に展開しています。このように鼻から脳への精密な神経配線が、多種多様な匂い分子の識別を可能にしています。一方、動物が匂いによって「快・不快」、「好き・嫌い」などの感情を抱いたり、過去の記憶を思い起こしたりするには、匂い情報が嗅球からさらに高次の嗅覚中枢へと伝わり、匂いの種類・質に応じて適切に処理される必要があります。しかし、嗅球から高次中枢への神経配線様式がどのようになっているのかについては、これまでほとんど分かっていませんでした。
魚類は、嗅覚系が発達しており、古くから嗅覚研究に適したモデル動物として、匂い受容メカニズムの解明に大きく寄与してきました。さらに、稚魚の身体が透明で、遺伝子工学的手法が確立している熱帯魚ゼブラフィッシュは、神経科学の分野で有用なモデル動物として注目されています。今回研究チームは、ゼブラフィッシュの嗅球の神経細胞(僧帽細胞)を蛍光可視化することに成功し、今まで知られていなかった新たな神経回路の存在を明らかにしました。
研究手法と成果
(1)嗅覚神経回路を可視化したトランスジェニックゼブラフィッシュの作製
研究チームは、2008年ノーベル化学賞(下村脩博士ら)の受賞対象となったクラゲ由来の緑色蛍光タンパク質(GFP)を用いて、嗅球の僧帽細胞を可視化したトランスジェニックゼブラフィッシュを作製しました。これまで、脳の中で嗅球の僧帽細胞だけにGFPを発現させることに成功した例はありませんでした。研究チームは、lhx2a※8という遺伝子のプロモーター領域(遺伝子の発現を調節するゲノムの領域)を利用することで、僧帽細胞特異的なGFPの発現に成功しました。また、神経線維やシナプス部位に集積する、ほかのタンパク質とGFPを融合させた「改変型GFP」を用いることで、僧帽細胞から高次中枢への神経配線パターンを詳細に解析することができるトランスジェニックゼブラフィッシュの作製にも成功しました(図2)。
(2)嗅球から手綱核への左右非対称な神経回路の発見
トランスジェニックゼブラフィッシュの解析から、ある糸球体クラスターと接続している僧帽細胞は、神経線維を終脳に投射するととともに、間脳※4の「手綱核」という部位にも投射することを発見しました。手綱核は、セロトニンやドーパミンなどを神経伝達物質として含有する神経細胞の活動を調節することで、感情の制御に深くかかわっていると考えられています。嗅球から手綱核への直接的な神経接続の存在は、これまでまったく知られていませんでした。また、嗅球から終脳への投射は左右対称ですが、手綱核への投射は、左右非対称であることが明らかになりました。すなわち、左と右のどちらの嗅球からも右の手綱核へ投射し、決して左には投射しないことが分かりました(図2、3、4)。
(3)単一僧帽細胞の遺伝子工学的標識法による匂い情報変換様式の解析
嗅球上に展開された「匂い地図」が、高次中枢へどのように伝達されているのかを解明するために、研究チームは、匂い情報の最小ユニット構造である個々の糸球体と、それに接続する個々の僧帽細胞の神経投射パターンを、単一細胞の遺伝子工学的標識法により解析しました。これは、研究チームが確立した、脳の中でたった一個の僧帽細胞を可視化したゼブラフィッシュを作製する技術で、このゼブラフィッシュを共焦点レーザー顕微鏡で観察すると、生きたままで特定の僧帽細胞の神経配線パターンが分かります。その結果、匂いの大まかな種類に対応した糸球体クラスターごとに、僧帽細胞が特有の投射パターンを持つことを明らかにしました(図5)。例えば、mGと名付けた糸球体クラスターと接続する僧帽細胞だけが、終脳と手綱核の両方に神経線維を投射し、ほかの糸球体クラスターからは終脳だけに投射します。一方、終脳内では、糸球体クラスターごとに一部重複しながら固有の投射領域を示す傾向がありました。これらの結果は、嗅球に入った匂い情報を、複数の脳領域に分配・再構成し、個々の脳領域で特定の情報を抽出することによって、匂いに応じた適切な行動が出力されることを示唆しています。
今後の期待
嗅覚は多くの動物にとって、食べ物の探索、危険の察知、社会性コミュニケーションや繁殖行動など、個体の生存と種の維持にかかわる重要な感覚センサーです。私たち人間にとっても、“匂い”は、快・不快の感情や過去の記憶を呼び起こしたり、味覚や食欲を左右したりする重要な役割を持っています。今回の研究で発見した嗅球から感情に深くかかわる手綱核への直接的な神経接続は、嗅覚と感情のつながりを探る糸口になると期待できます。また、脳の左右非対称性は、左右の脳が機能分担することで情報処理を効率化しているという考えとともに、非対称性の方向が種ごとに共通することで、社会行動に協調性が生じて、種の保存に有利に働くという説が提唱されています。例えば、群れをなして行動する魚では、外敵に対する危険回避の際に、同一種の個々の魚は、同じ方向にターンする傾向があることが知られています。今回発見した「嗅球-手綱核」間の神経回路は、左右非対称で、その方向性が一定なことから、このような動物行動の左右非対称性に関与する可能性が考えられます。今後、この神経回路の解析を通じて、「情動の発現」や「動物行動の左右非対称性」の基盤となる神経回路メカニズムが解明されるものと期待できます。
発表者
理化学研究所
脳科学総合研究センター シナプス分子機構研究チーム
チームリーダー 吉原 良浩(よしはら よしひろ)
Tel: 048-467-1699 / Fax: 048-467-2306
副チームリーダー 宮坂 信彦(みやさか のぶひこ)
Tel: 048-467-9674 / Fax: 048-467-9689
お問い合わせ先
脳科学研究推進部 鈴木 一郎(すずき いちろう)Tel: 048-467-9654 / Fax: 048-462-4914
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.ゼブラフィッシュ
学名:Danio rerio。インド原産の体長5センチメートルほどの小型熱帯魚。飼育が容易で多産。稚魚の体は透明なので、体の内部の発達過程を生きたままで観察することができる。遺伝子工学的な手法を用いることで、特定の神経細胞を蛍光タンパク質によって可視化したり、特定の遺伝子の機能を阻害したりすることが可能。
- 2.嗅球
嗅覚の一次中枢として機能する脳の領域。嗅細胞の神経線維が直接接続している。 - 3.僧帽細胞
嗅球の糸球体に入力された匂いの情報を受け取り、高次中枢へと伝える嗅球の神経細胞。細胞の形が「司教の冠(miter)」と似ていることからこの名が付けられた。一つの僧帽細胞は一つの糸球体から匂い情報を受け取る。 - 4.終脳、間脳
脳の領域の名称。前方から終脳、間脳、中脳、後脳、髄脳。嗅球は、終脳の最も前方に位置する。 - 5.手綱核
終脳と中脳を結ぶ伝達経路を構成する脳の領域。セロトニン作動性およびドーパミン作動性神経の活動調節に関与すると考えられている。近年、ゼブラフィッシュを用いた解析から、遺伝子発現、細胞構成、神経配線などに左右非対称性があることが明らかとなっている。 - 6.嗅覚受容体
鼻腔に入ってきた匂い分子を認識する受容体。鼻の奥に存在する嗅細胞に発現している。多種多様な「匂い分子」に対応できるように、マウスでは約1,300種類、ヒトでは約350種類、魚では約200種類がゲノム上に存在している。 - 7.糸球体
嗅球の表面に並んだ神経線維からなる球状の構造体。一つの糸球体は、同じ嗅覚受容体を発現する嗅細胞に神経支配されており、その受容体と結合する匂い分子の情報を表現している。 - 8.lhx2a
遺伝子の発現を調節する「転写因子」の一種。マウスでは、嗅覚受容体遺伝子の発現を誘導することが知られているが、ゼブラフィッシュでの機能は不明。
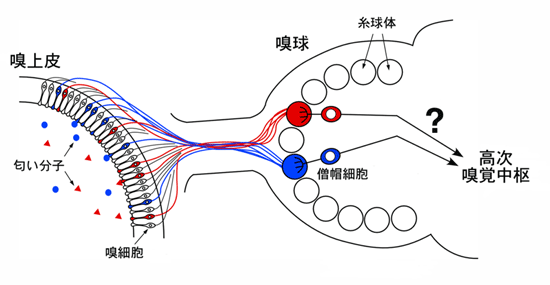
図1 嗅覚神経回路の構造
鼻腔に入った匂い分子(●、▲)は、鼻の奥にある嗅上皮で嗅細胞によって受け取られる。個々の嗅細胞はたった一種類の嗅覚受容体を発現し、特定の化学構造を持った匂い分子の情報を脳へと伝える。同じ受容体を発現した嗅細胞(赤や青)は、神経線維を嗅球の同じ糸球体に投射する。糸球体で嗅細胞から入力を受け取った僧帽細胞は、その情報をさらに高次中枢へと伝える。
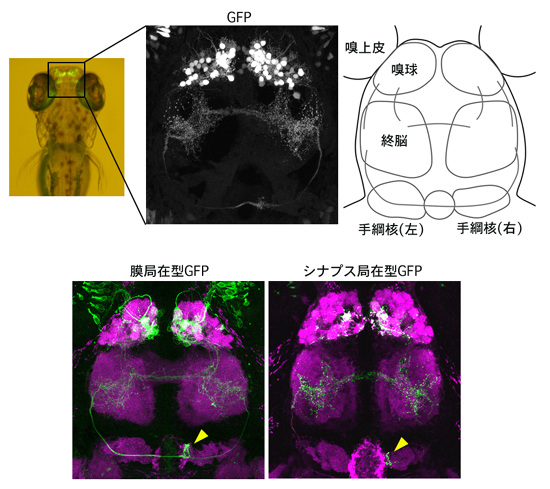
図2 僧帽細胞をGFPで可視化したトランスジェニックゼブラフィッシュの稚魚
受精後7日目の稚魚の脳を背側から観察。脳内では、嗅球の僧帽細胞だけをGFPで可視化している(上段)。改変型GFP(下段、緑)を用いることで、神経線維の投射パターン(左)およびシナプス部位(右)を明瞭にラベルすることができる。免疫組織化学により脳の構造を可視化した(紫)。僧帽細胞は右手綱核に投射するが(下段、黄色の矢尻)、左手綱核には投射しない。
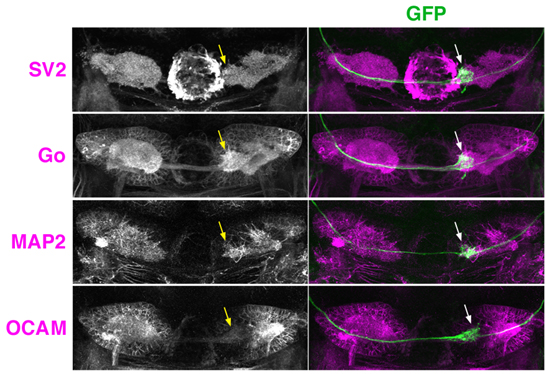
図3 嗅球から手綱核への非対称な投射
稚魚の手綱核をさまざまなマーカー抗体を用いて染色した画像(左パネルの白および右パネルの紫)。嗅球の僧帽細胞に由来する神経線維(右パネルの緑)は、右手綱核の内側部(矢印)に特異的に投射している。
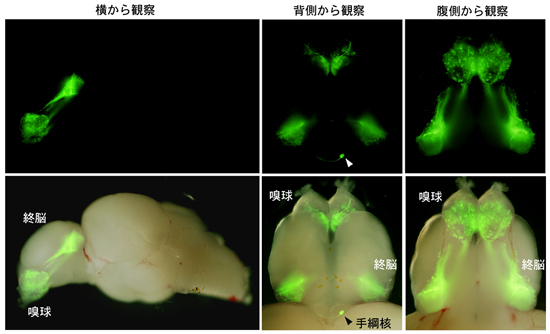
図4 僧帽細胞の右手綱核への投射は成魚でも維持されている
上段:蛍光画像 下段:蛍光画像と通常の画像の重ね合わせ
トランスジェニックゼブラフィッシュの成魚の脳を横(左パネル)、背側(中央パネル)、腹側(右パネル)からそれぞれ観察した。嗅球から右手綱核への投射(中央、矢尻)は成魚でも維持されている。
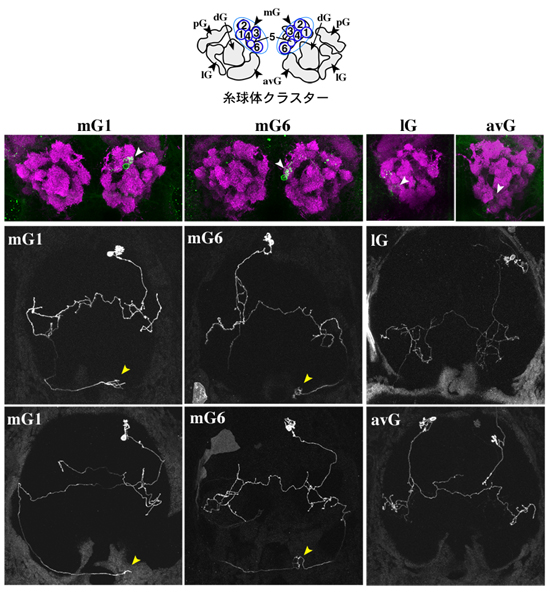
図5 単一僧帽細胞の遺伝子工学的標識法による匂い情報伝達様式の解析
糸球体のmGクラスターと接続する僧帽細胞の多くは、左右の終脳と右の手綱核に投射する(左および中央パネル、黄色の矢尻)。一方、ほかの糸球体クラスター(lGやavGなど)と接続する僧帽細胞は終脳に投射するが、手綱核には投射しない。