ポイント
- 陸上植物に共通する葉緑体分裂装置の構成因子の量が、分裂の速度を決定
- 葉などへの組織分化に伴って、葉緑体の数や大きさを変える機構も存在
- 葉緑体の数や大きさを自在に変化させた新植物の開発が可能に
要旨
独立行政法人理化学研究所(野依良治理事長)は、植物の葉緑体分裂装置※1の構成因子であるPDV1、PDV2(PLASTID DIVISION 1と2)※2タンパク質の量によって葉緑体の分裂速度が変化し、葉緑体の数や大きさが調節されていることを発見しました。これは理研基幹研究所(玉尾皓平所長)宮城島独立主幹研究ユニットの岡﨑久美子ユニット研究員、壁谷如洋ユニット研究員らと、理研植物科学研究センター(篠崎一雄センター長)植物ゲノム機能研究グループの松井南グループディレクター、市川尚斉研究員らによる共同研究の成果です。
葉緑体は、光合成、アミノ酸合成、脂質合成などの重要な機能を担う植物の細胞内小器官です。葉緑体は新たに合成されることはなく、既存の葉緑体の分裂でしか、数を増やすことができません。研究グループは、葉緑体分裂装置の構成因子であるPDV1、PDV2の量を人為的に増やすと葉緑体数が増えて大きさが小さくなり、PDV1、PDV2の量を減らすと逆に葉緑体数が減って大きさが大きくなる、という現象を発見しました。つまり、PDVが葉緑体の分裂速度を制御していることを明らかにしました。さらに、PDVの量が植物ホルモンであるサイトカイニン※3によって制御され、組織ごとに、葉緑体分裂の速度が変化していることを明らかにしました。この仕組みを原始的な陸上植物であるコケ植物のヒメツリガネゴケと、高等な種子植物のシロイヌナズナで観察したことから、すべての陸上植物共通の機構であることが強く示唆されました。
この成果は、葉緑体分裂の制御機構を初めて明らかにしたもので、植物の成長の基本的な仕組みの理解に貢献します。また、葉緑体の数や大きさを自在に変化させて、デンプン粒の大きさを変えた作物、強光に強い作物、光合成能を高めた作物などの開発が期待されます。
本研究成果は、米国の科学雑誌『The Plant Cell』オンライン版(6月30日付け:日本時間7月1日)に掲載されます。
背景
葉緑体は光合成を行う細胞内小器官で、光合成は植物を含めたすべての生命を支えているといっても過言ではありません。葉緑体は、今から10~20億年前に、光合成を行う独立した生物(シアノバクテリア)が、植物の祖先細胞に取り込まれてできたと考えられています。そのため、もはや植物細胞の一部となった今でも、ゼロから作り出すことはできず、葉緑体自身が分裂することによってのみ増えることができます。
葉緑体の分裂は通常、中央が徐々にくびれ、2つにちぎれることで完了します。このくびれ部分には、数種類のタンパク質からなる葉緑体分裂装置がリング状に存在し、その直径がだんだん狭まることによって分裂が進行します(図1)。分裂装置を構成するタンパク質には、葉緑体の祖先となったシアノバクテリアがもともと分裂に使っていた先天的なものと、それを取り込んだ植物細胞が分裂をコントロールするために新たに付け加えた後天的なものがあることが分かっています。先行研究で、宮城島進也独立主幹研究員はPDV1とPDV2(PLASTID DIVISION 1と2)というタンパク質が陸上植物特異的な葉緑体分裂装置の構成因子であり、後天的に付け加えられたタンパク質であることを明らかにしました(Miyagishima et al. Plant Cell, 2006)。しかし、このPDV1とPDV2が、藻類※4が陸上植物へと進化する過程で葉緑体分裂装置に新たに加えられた意義については謎でした。
藻類の細胞あたりの葉緑体の数はほとんど一定で、その分裂は細胞自体の分裂に合わせて一定のペースで起きています。一方、陸上植物では、細胞によって葉緑体の数や大きさ、分裂のペースは大きく違います。新しい葉が次々と作られている分裂組織の付近では、葉緑体は小さく活発に分裂していますが、成長した大きな葉では、葉緑体は大きく分裂は活発ではありません。藻類に比べて構造が複雑な陸上植物では、葉の成長などに合わせた葉緑体分裂の調節が必要です。しかし、葉緑体分裂の制御の仕組みはほとんど分かっていませんでした。今回、PDV1、2に着目し、葉緑体分裂の調節機構を解明することを目的に研究を行いました。特に、陸上植物で新たに進化した植物の成長に合わせて葉緑体の分裂をコントロールする仕組みの解明を目指しました。
研究手法と成果
研究グループは、種子植物であるシロイヌナズナを使って、植物ゲノム機能研究グループの開発したFOXハンティングシステム※5により、本葉※6の細胞中の葉緑体の分裂を促進させる遺伝子を探索しました。その結果、葉緑体分裂装置の構成因子である、PDV2遺伝子を過剰に発現させると葉緑体の分裂が促進することが分かりました。PDV2と共に働くと考えられているPDV1も同様に過剰発現させたところ、同様に分裂を促進することが分かりました(図2)。PDVの量を減らすと葉緑体の分裂頻度が低下することや、ほかの構成因子を過剰に発現させても分裂は促進せず、かえって阻害することもあることなどから、PDVの量が葉緑体の分裂の頻度を決定していると考えました。
植物自身がPDVの量を調節することで葉緑体の分裂を制御しているかを調べるために、シロイヌナズナの植物体をばらばらにして葉の大きさごとに分け、その中に含まれるタンパク質の量を調べました。するとPDVの量は、葉緑体分裂の盛んな分裂組織や若い未熟な葉で多く、大きく成長した葉では少ないことが分かりました(図3)。ほかの構成因子はそのようなパターンを示さなかったことから、植物は葉の成長に伴ってPDVの量を減らし、葉緑体の分裂を調節するという仕組みを持っていることが示唆されました。
FOXハンティングシステムによるさらなる解析で、CRF2遺伝子を過剰発現させた場合にも葉緑体の分裂は促進し、このときPDV2の発現が上昇していることが分かりました。CRF2タンパク質は、成長や細胞分化を制御する植物ホルモンの一種であるサイトカイニンに応答する転写因子です。植物に外からサイトカイニンを与えてみると、CRF2の発現上昇に伴ってPDV2の発現が上昇し、葉緑体の分裂を加速することが分かりました。このことから、PDVはサイトカイニンによって制御される細胞分化のプログラムに従って、葉緑体の分裂ペースを調節する役割を担っていると分かりました。
PDVは陸上植物に広く共通して存在します。その進化的側面を知るために、陸上植物の祖先から最初に分岐したと考えられている、コケ植物のヒメツリガネゴケを使った実験を行いました。ヒメツリガネゴケでPDV遺伝子の1つを過剰発現させたところ、種子植物のシロイヌナズナのときと同様に、葉緑体分裂の促進が起こりました。また、ヒメツリガネゴケの原糸体※7にサイトカイニンを与えると、PDV遺伝子の発現が上昇し、サイトカイニンによって分化が誘導された細胞では葉緑体分裂が加速しました。これらのことから、PDVの機能や役割は、陸上植物に共通であることが示唆されました。約5億年前に陸上植物の祖先が上陸する際、PDV遺伝子を獲得したことにより、葉緑体分裂を調整できるようになり、葉緑体の数と大きさの制御が可能になったものと考えられます(図4)。その結果、植物の細胞はより多様に分化して、複雑な構造に進化できるようになり、地球全体に広がるほどの繁栄が植物にもたらされたことが想像できます。
今後の期待
今回の研究結果により、葉緑体の分裂制御と細胞分化の結びつきが初めて遺伝子レベルで明らかとなりました。葉緑体の分裂という細胞内での現象と、細胞分化という植物全体での現象の関係がつながったことは、植物の成長の基本的な仕組みを理解する上で重要です。
デンプン粒の大きさは、葉緑体の大きさに依存することが知られています。今回、PDV1、PDV2の量を人工的に変化させることで、葉緑体の数や大きさを変化させることができました。この成果を利用して、例えば、作物に含まれるデンプン粒の大きさを変えることが可能かもしれません。デンプンの大きさを変えて新しい食感を得たり、工業的に利用価値の高い品種の開発につながる可能性があります。また葉緑体は、適切な量の光を得るためや、活性酸素を発生するような強光から逃れるために、光量に応じて細胞内を移動することが知られています。葉緑体の数と大きさを人工的に変化させることで、この運動効率を変え、強光に強い植物を作出できる可能性も見えてきました。
発表者
理化学研究所
基幹研究所 宮城島独立主幹研究ユニット
ユニット研究員 岡﨑 久美子(おかざき くみこ)
ユニット研究員 壁谷 如洋(かべや ゆきひろ)
Tel: 048-467-9332 / Fax: 048-467-9329
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.葉緑体分裂装置
分裂中の葉緑体のくびれの部分に見られるリング状のタンパク質複合体。このリングが収縮することで、葉緑体が2つに分断される。葉緑体の2枚の膜の内側、中間、外側にまたがっている。葉緑体の祖先のシアノバクテリアが細胞分裂に使っていた因子と、植物細胞が分裂を制御するために新たに付け加えた因子とのキメラ構造である。 - 2.PDV1、PDV2(PLASTID DIVISION 1、2)
陸上植物の葉緑体分裂装置の構成因子となるタンパク質。葉緑体の外側の膜(外包膜)を貫通している。分裂装置の構成因子の1つであるダイナミンタンパク質を分裂装置へ誘導するのに必要であることが知られている。 - 3.サイトカイニン
植物ホルモンの一種。細胞分裂の促進、細胞周期の調節、老化阻害、腋芽の活性化など多様な生理活性を持つ。アデニンの6位の窒素原子に炭素5つのプレニル基を持つ構造が基本骨格であり、側鎖構造の違いによりトランスゼアチン〔tZ〕、イソペンテニルアデニン〔iP〕などが知られている。中でもtZの生理活性が最も強い。植物のほかにアグロバクテリウムなどの一部の土壌細菌、キイロタマホコリカビなどの粘菌が合成する能力を持つことが知られている。 - 4.藻類
酸素発生型光合成を行う生物の中から、コケ植物、シダ植物、および種子植物を除いた生物。ほとんどは水中または湿潤なところに生息する。構造は単純で、根、茎、葉といった構造を持たない。 - 5.FOXハンティングシステム
ホスト生物に目的生物の完全長cDNAを導入、高発現させた形質転換体を作製し、ある特定の条件で選別して、最終的にその個体から導入した遺伝子を取り出すというゲノムワイドな有用遺伝子探索技術。 - 6.本葉
種子植物の子葉の後に出てくる葉。 - 7.原糸体
コケ植物で、胞子が発芽してできる糸状の配偶体。
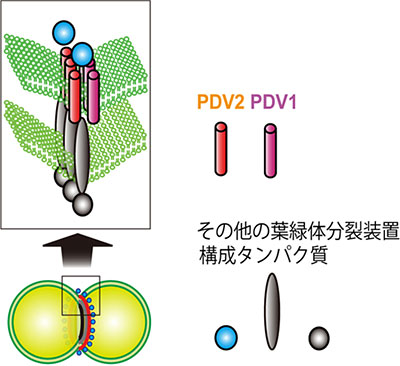
図1 葉緑体分裂装置の構造
葉緑体は分裂面にリング状に存在する葉緑体分裂装置によって中央がくびれ、2つに分かれる。分裂装置は複数種類のタンパク質からなり、その一部であるPDV1、PDV2は外側の膜(外包膜)に局在している。
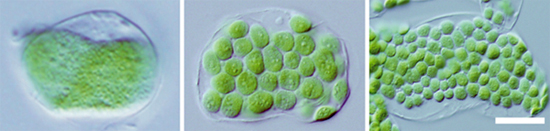
図2 PDV1、PDV2の量を変えたときの葉緑体の様子
左:PDV1とPDV2が発現しない植物の細胞
中央:正常な植物(野生型)の細胞
右:PDV1とPDV2が過剰に発現している植物の細胞
スケールバー=10マイクロメートル
種まきから3週間後の植物の葉を切り取り、ばらばらにして細胞を固定し顕微鏡で観察した。
PDV1、PDV2が共に発現していない植物では、葉緑体の分裂が少なくなり、細胞内に巨大な葉緑体が1つだけ観察された(左)。逆にPDV1とPDV2が過剰に発現すると、細胞当たりの葉緑体の数は正常な植物の場合の(中央)ほぼ2倍になり、大きさは小さくなった(右)。
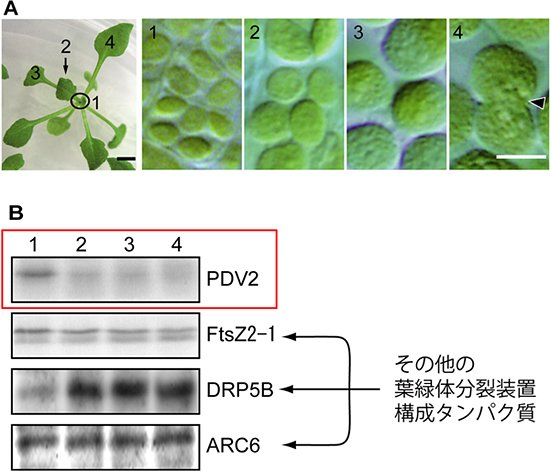
図3 葉の発達段階の違いによる葉緑体の変化と、そのときのPDV2などの葉緑体分裂装置構成タンパク質の量の変化
A:シロイヌナズナの葉の発達と葉緑体の変化
植物体の中心部分(左写真の1)では新しい葉がつくられている。新しくできた葉の細胞の中では小さな葉緑体が盛んに分裂している(右写真1)。左写真2→3→4と葉が成長するにつれて、葉緑体の分裂は少なくなり、大きさが大きくなる(右写真2~4)。大きく成長した葉でも分裂は起きているが(右写真4矢印)、その頻度は低い。 スケールバー=2ミリメートル(左写真)、5マイクロメートル(右写真1~4)
B:葉の発達段階でのPDV2とそのほかの葉緑体分裂装置構成タンパク質の量の変化
Aの1~4の段階の葉をそれぞれ集めて、そこに含まれるタンパク質の量を調べた。PDV2は1の段階で多く、葉が成長してきた2の段階ではすでにかなり減っていた。そのほかのタンパク質(FtsZ2-1、DRP5B、ARC6)はずっと一定であるか、成長した葉の方が量は多かった。
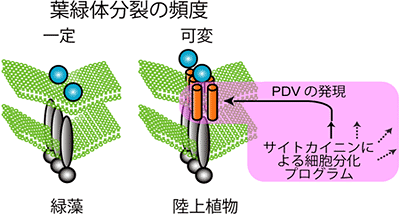
図4 陸上植物で進化したPDVによる葉緑体分裂のコントロール機構
緑藻は細胞分裂に合わせて、ほぼ一定の速度で葉緑体分裂をする。その葉緑体分裂装置にはPDVはない。従って、陸上植物の祖先がPDVを葉緑体分裂装置の中に取り入れたと考えられる。PDVの量は、植物ホルモンであるサイトカイニンによって起きる細胞分化プログラムによって変化する。PDVによって陸上植物は葉緑体分裂の頻度をコントロールできるようになり、細胞の葉緑体の数や大きさを変化させることができるようになった。