2009年7月10日
独立行政法人 理化学研究所
カメが甲羅を作った独特の進化過程を解明
―カメは体壁を折り曲げ、肋骨を広げて甲羅を作る―
ポイント
- カメの特異性は胚発生後期に現れ、発生前~中期の基本的発生過程は他の羊膜類と同様
- カメは祖先の後期発生プログラムの変更により、甲羅を得たと推測
- カメの発生過程から得た進化シナリオは、原始的カメの化石記録とも一致
要旨
独立行政法人理化学研究所(野依良治理事長)は、羊膜類※1の中でも甲羅などの独特の骨格形態を持っているカメの形態がどのようにして得られたのか、という謎に挑み、カメの発生後期過程において腹側の体壁が内側に折れ込むことによって骨格の位置関係が変わる様子を観察し、カメの進化を発生学的に明らかにしました。これは、発生・再生科学総合研究センター(竹市雅俊センター長)形態進化研究グループの倉谷滋グループディレクター、長島寛研究員らによる成果です。
カメは、羊膜類の中でも特徴的な形態進化を経てユニークな骨格パターンを作り出しています。カメの甲羅は背側の甲羅(背甲)と腹側の甲羅(腹甲)からなり、そのうち背甲は肋骨が扇状に広がって形成されたもので、これに伴い周辺の骨格も変化しています。特に、は虫類や鳥類、ほ乳類を含む一般の羊膜類では、肩甲骨が肋骨の外側にあるのに対し、カメの肩甲骨は肋骨である背甲の内側にあり、肩甲骨と肋骨が逆転した位置関係を示しています。この逆転がいかにして起きたのか、カメと鳥類やほ乳類の胚の中での肩甲骨や肋骨の発生過程を比較し、原始的なは虫類からカメが進化した際の発生過程の変化を推定しました。
その結果、カメの肩甲骨は胚発生の中期までは、ほかの羊膜類と同様に肋骨の外側前方にありますが、胚発生後期に腹側の体壁が内側に折れ込むため、肩甲骨が二次的に肋骨の下へと移動し、肋骨の内側に位置するように見えることが分かりました。さらに興味深いことに、このカメの発生中期の胚形態は2008年11月に発表された、最古の化石カメと見なされるOdontochelys(オドントケリス;中国の三畳紀後期2億2千万年前の地層から発見、Liら、Nature、2008)の形態と非常によく似ていました。つまりOdontochelysには背甲がまだなく、その肋骨も扇状に広がらず、発生中期のカメの胚に見るように、肩甲骨が肋骨の外側前方にありました。おそらく現在のカメは、Odontochelysのような祖先動物の後期発生プログラムをさらに変更し、肋骨を扇状に広げることで、肩甲骨を覆う背甲を得たと考えられます。本研究成果は、米国の科学雑誌『Science』(7月10日号)に掲載されるに先立ち、オンライン版(7月9日付け:日本時間7月10日)に掲載予定です。
背景
カメの甲羅は背甲と腹甲からなり、そのうち背甲は脊椎を中心に肋骨が扇状に横方向に開いてできたものです。この独特の骨格形成に伴って、カメでは骨や筋肉などの体のいくつかのパーツの位置関係が、羊膜類の中で独特なものとなっています。とりわけ、カメの肩甲骨は肋骨の内側にあり、肩甲骨が肋骨の外側にあるヒトを含めた一般的な羊膜類に比べて、その位置関係が逆転しています(図1)。同様に肩や二の腕を胴体につないでいる筋肉もカメでは甲羅の内側にあり、これも一般的な羊膜類での位置関係とは異なっています(図2)。このようにカメは羊膜類の中で非常に奇妙な形態を示しています。しかし、これらの特徴も、肋骨の外側に肩甲骨や筋肉がある、一般的な形態を持った祖先動物が変化してできたと考えられます。
カメの進化は、劇的な進化の例として有名です。たとえばコウモリや鳥の翼は進化の過程で1つ1つの前足の骨が伸びたもので、骨や筋の位置関係は元のまま維持しています(図3)。ところが、カメの甲羅、つまり肋骨と肩甲骨の位置関係はほかの羊膜類のものとは合致せず、羊膜類の中で進化上保存されてきた骨格・筋肉の位置関係のパターン(形態学的相同性※2)が、カメにおいてだけ崩れているように見えます。どうやら、カメの進化では、何か形態発生上の質的な変化が生じているようです。このように、カメの肩甲骨や肋骨が示している独特の進化パターンは、100年以上にわたり、解剖学、形態学、古生物学の大きな謎とされてきました。
動物の形は発生過程を経て作られ、異なった形の動物には異なった発生過程があります。進化は「形の変化」の歴史であり、すなわち「発生過程の変化」の歴史と言い換えることができます。
そこで研究グループでは、どのようにしてカメのようなユニークな形態が進化上できたのかを理解するため、カメの胚発生に着目し、解析を進めました。
研究手法
カメの進化過程において生じた発生プログラムの変化を探るためには、変化する前の発生過程、すなわちカメの祖先動物(化石)の発生過程を理解しなければなりません。しかし、発生過程は化石としては残りません。そこで現生のカメやカメ以外の羊膜類の発生を比較することによって、カメの祖先の発生過程を推定しました。近年の分子系統学的研究(動物間のDNA配列の比較)は、カメ類と鳥類が互いに近い系統に属していることを示しています(図4)。そこでカメの一種であるスッポン(甲羅表面の角質がないカメ)と鳥類のニワトリ、さらに、カメとニワトリの分岐以前に原始的なは虫類から分かれたと考えられている、ほ乳類のハツカネズミを比較対象としました。すなわち、カメ、ニワトリ、ハツカネズミの発生過程を比較したとき、ニワトリとハツカネズミに共通する形質が見つかれば、それはおそらく羊膜類の共通祖先がすでに持っていた発生プログラムだろうと考えることができます。一方、カメだけにある形質はおそらく、カメの祖先動物がカメに進化する際に起こった発生プログラムの変化を示していると考えられます。
このような考えのもとに、X線CTスキャンや組織切片の再構築※3、マーカー遺伝子の発現パターン、免疫組織化学染色※4を用いて、カメ、ニワトリ、ハツカネズミの発生過程の、いつ、どこに、どのような位置関係で、骨格や筋ができてくるかを比較・観察しました。
研究成果
(1)発生中期までは、カメ、ニワトリ、ハツカネズミは共通の形態発生
胚発生の中期までは、カメ、ニワトリ、ハツカネズミのすべてで、共通したパターンをもとに肩甲骨、肋骨、筋肉が発生していました。すなわち、カメの肩甲骨だけでなく、肩と体幹をつなぐ筋も、肋骨と肋間筋の原基(筋板)の外側に見られ、この位置はほかの動物で見られるものと同じでした。ただし、カメの肋骨はほかの動物のものと比べると相対的に短く、背側にとどまっていました(図5)。
(2)カメでは胚発生後期に体壁が内側に折れ込まれ、肩甲骨が移動する
発生後期、カメの肋骨は横方向に伸び、扇状に広がって肩甲骨に後背側から覆いかぶさり、成体のカメで見られるような骨格の位置関係ができることを確認しました(図1、5)。このカメ独特の肋骨の広がりは、肋骨の先端近傍にあるCR (Carapacial Ridge:甲稜=こうりょう)という、将来の甲羅の縁となる部分によって引き起こされることが分かっています(長島ら、Development、2007)(図6)。CRによって肋骨が扇状に広がるのに伴い、腹側の筋板は肩甲骨や付随する筋を含め、内側に大きく折れ曲がっていきます。すなわちカメは、筋板を含む体壁を内側に折り込み、肩を体の内側に引き込むことで、見かけ上の肋骨と肩の位置関係を逆転させていたわけです(図1、5、7)。
肩の筋肉の1つである前鋸筋(ぜんきょきん)は、この折れ込みの前、すなわち発生中期には肩甲骨と肋骨を結び、肋骨の外側にありました(図8)。この形は発生中期と成体のニワトリやハツカネズミと同じでした。筋は、その後も肩甲骨、肋骨とのつながりを維持するので、折れ込みに伴い、肋骨の内側へとその位置を変えることになったのです(図6、8)。つまり、見かけ上の位置が大きく変わっても、筋の骨格との位置関係は実は変化していないのです。同様のことは、肩甲骨と脊椎骨をつなぐ肩甲挙筋や菱形筋についても確かめることができました。
一方でカメには、骨に付着する位置を大きく変えた筋もあります。たとえば、ニワトリ、ハツカネズミの広背筋は、肩関節の後ろに広がって二の腕と胴体を結んでいますが、カメでは肩関節よりも前側に発生しました(図2、9)。また、ニワトリ、ハツカネズミの胸筋は、体の浅い所にあって胸骨(胸の正中にある骨)に外側からくっついていますが、カメの胸筋は腹甲に内側から付いています。前鋸筋とは異なり、これらの筋肉は発生中期には、まだ胴体側の骨格へ付着していない、発生の遅い筋であることが分かりました。
(3)カメの進化は、「折れ曲がり」と「新しい結合」によって説明できる
肩の筋は、発生の早期に骨格とのつながりが達成される前鋸筋、肩甲挙筋、菱形筋(体壁筋)と、発生後期に作られる広背筋、胸筋など(前足の筋肉)の2種類に分けられます。前者の筋は、発生初期から周囲の骨格との位置関係を変えることなく発生するもので、その結果として骨格とのつながり方は変わらず、カメの発生では大きく折れ曲がります。一方、後者の筋のもととなる細胞は、Lbx-1遺伝子※5という発生調節遺伝子を発現していました(図9)。この遺伝子は、筋のもとになる細胞が移動する時に働く遺伝子です。これらの細胞は発生初期にまず、胴体から腕の中にいったん移動し、その後、発生後期に胴体へと伸び出し、胴体への付着点を作ります。カメでは、この胴体へ戻る過程でこれらの筋が新たな付着点を作り出しているわけです。言い換えれば、この発生プログラムは、進化的に変化しやすい部分であると想像できます。カメの体は、発生プログラムのなかで変化可能な部分をうまく使い、もっぱら「体壁の折れ曲がり」と、「筋の新しい付着点の形成」によってもたらされているといえます。
(4)化石記録との一致
非常に興味深いことに、2008年11月に発表された化石動物Odontochelysの形態は、今回観察したカメの発生中期の姿に非常によく似ています(図10)。Odontochelysには、腹甲はありましたが背甲はなく、現在のカメの祖先になった動物であったという可能性が考えられています。Odontochelysの肋骨は、現在のカメと同様に横に広がっているものの、扇状にはなっておらず、肩甲骨が肋骨の前外側に確認され、カメの胚の発生中期の形態とそっくりなのです。つまり、Odontochelysの発生過程ではCRのようなものはあったかもしれませんが、それは扇状の肋骨の広がりを導く能力はなかったらしいと推察できます。おそらくカメの進化は、このような動物の後期発生過程を変更し、CRの能力を強化して、背甲を完成させることでなされたのではないかと考えられるのです。
今後の期待
本研究により動物の形には、変化させやすい場所とさせにくい場所があることが明らかとなりました。ユニークな体を持った動物が存在しているからには、それを作り出した進化的な変化の歴史があるはずで、それは正しい方法で研究すると細胞や遺伝子の働きをベースにした、発生プログラムの変更として理解できるはずです。とりわけ今回の研究では、遺伝子や細胞レベルでの発生学の知見と、解剖学の情報を密に組み合わせ、さらに古生物学的な情報の中にその証拠の一端を見いだそうという方法が成功したのだと思います。私たち人間の体の解剖学的なパターンも、発生プログラムの結果だという見方が必要です。カメの研究は、ヒトの体とかなり同じデザインを持ちながらも、発生プログラムにおける数カ所の変化で、ユニークな形態を作りうる可能性を垣間見せてくれました。「ゲノムから形へ」という、発生という現象の性質を理解するケーススタディとして、研究グループのカメ研究が1つのヒントになるのではないかと考えます。
発表者
理化学研究所
発生・再生科学総合研究センター形態進化研究グループ
研究員 長島 寛(ながしま ひろし)
Tel: 078-306-3064 / Fax: 078-306-3370
お問い合わせ先
発生・再生科学総合研究センター広報国際化室中込 咲綾(なかごみ さや)
Tel: 078-306-3310 / Fax: 078-306-3090
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.羊膜類
脊椎動物のうち、胚が羊膜に包まれているものの総称。は虫類、鳥類、ほ乳類を指す。一部を除き生活環の全体を通じて陸上で生活する。 図4参照。 - 2.形態学的相同性
骨や筋肉などの解剖学的単位が、異種類の生物間で同一の配列を示し、その構造に何らかの共通点を持つこと。例えば、さまざまな動物で前足の骨を比べたとき、肩甲骨-二の腕の骨-腕の骨-手の骨のように、動物間で大まかに同じ形をした骨が同じ順序で並んでいる。このとき、動物間で対応するそれぞれの骨の間の関係性のことを「形態学的相同性がある」、また動物間の前足全体としても互いに「形態学的相同性がある」という。形態学的相同性が現れる原因は、これらの動物が同じ祖先を共有していることに由来する。 図3参照。 - 3.組織切片の再構築
胚は小さく、組織が十分に出来上がっていないため、骨や筋肉を成体のように解剖して調べることができない。そこで、胚を6μm程度の薄切りにし、骨や筋肉を染色して写真撮影し、これらの写真を再びコンピュータで集めて3次元的に復元すると、胚における骨や筋肉の位置や形が分かる。 - 4.免疫組織化学染色
胚の中にある形成途上の筋肉や骨を、その周囲の組織から見分けることは容易ではない。そこで、あらかじめ茶色い色素をつけた抗体の水溶液に胚を漬けることで、それらを色素で染め出すことができる。 - 5.Lbx-1遺伝子
筋肉ができる際には、胚の中で将来筋肉細胞になるもとの細胞ができる場所と、成体で筋肉がある場所が異なる場合がある。例えば筋板をつくる筋肉は、胚の背側にあった筋肉のもとの細胞が腹側に移動したものであるし、四肢の筋肉も背側の細胞が四肢の中に移動してきてできるのである。しかしこれらは移動の方式が異なっている。前者は細胞同士が密に固まってシート状に移動し、 Lbx-1という遺伝子を発現しない。一方で後者では細胞がばらばらになって移動し、移動し始めてからその後筋肉ができるまでの間、 Lbx-1を発現するのである。 Lbx-1遺伝子は、移動の際に細胞がその通り道を探すのに役立つ別の遺伝子の発現を促すものであることは分かっているが、その標的遺伝子はまだ分かっていない。
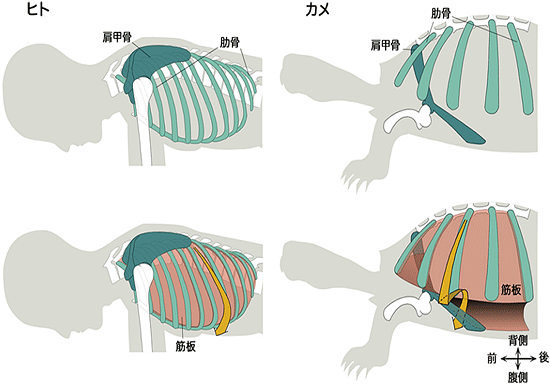
図1 ヒトとカメの肋骨と肩甲骨の比較
(上)ヒトとカメの骨格の比較。ヒトの肩甲骨は肋骨の外側にあるが、カメの肩甲骨は肋骨の内側にある。
(下)骨格に肋間筋の原基である筋板を追加した。筋板(赤)はヒトでは背側から腹側へと肋骨の間に沿っているが(矢印)、カメでは肋骨先端より腹側では、内側へ折れ曲がり、肩甲骨の内側を通っている(矢印)。
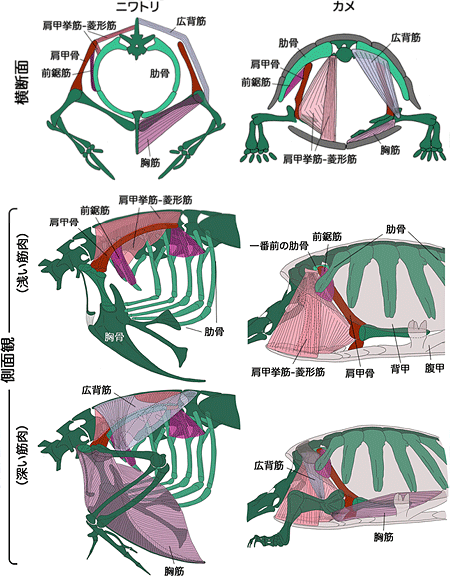
図2 ニワトリとカメの肩、二の腕の筋肉の比較
ニワトリの肩、二の腕の筋肉はすべて肋骨の外側にある。この位置関係はヒトも同じで、一般的な羊膜類に共通のものである。カメではこれらすべての筋肉が甲羅の内側にある。広背筋は、ニワトリでは肩関節の後ろ、背中に広がっているが、カメは肩関節の前にある。ニワトリの胸筋は胸骨を外側から覆うが、カメの胸筋は腹甲に内側から付いている。
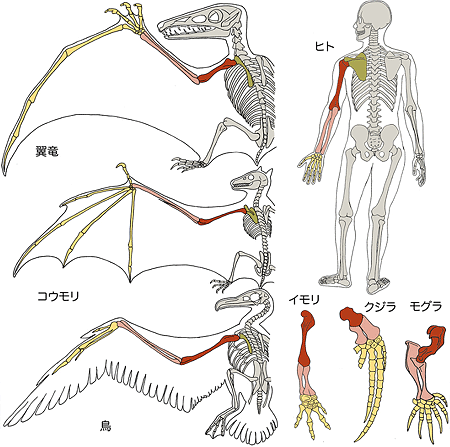
図3 前足の相同性
さまざまな動物の前足の骨格。緑は肩甲骨、赤は二の腕の骨である上腕骨(じょうわんこつ)、ピンクは腕の骨である橈骨(とうこつ)と尺骨(しゃっこつ)、黄色は狭義の手の骨である手根骨(しゅこんこつ)、中手骨(ちゅうしゅこつ)、指骨(しこつ)を示す。前足の形は動物の生活様式に合わせてさまざまに変化しているが、骨同士のつながり方は同じである。
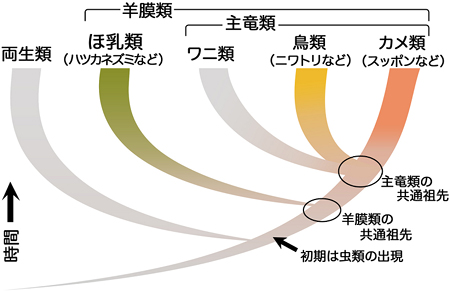
図4 脊椎動物の系統
水棲生活から完全に離れた両生類から原始的なは虫類が進化した。その後、初期は虫類の中からほ乳類が分かれ、残ったは虫類からさらにいくつかのグループが分かれた。その1つの主竜類には、カメ類、鳥類(鳥類は特殊化したは虫類)、ワニ類が含まれ、これらは共通の祖先から進化したと考えられている。主竜類とほ乳類は、羊膜類というグループに含まれ、原始的なは虫類を共通の祖先として持つ。
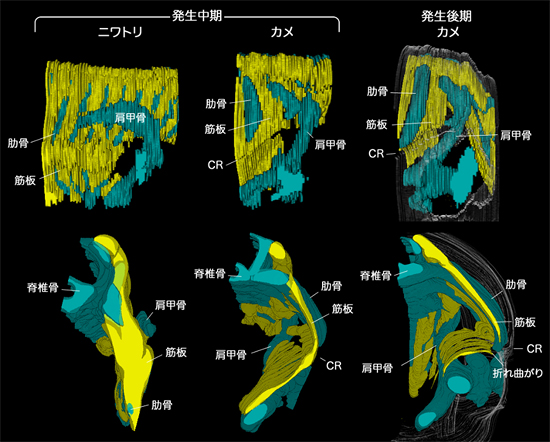
図5 カメにおける体壁の折れ曲がり
発生中期のニワトリ胚(左)とカメ胚(真ん中)、および発生後期のカメ胚(右)の組織切片の再構築画像。上は側面観(右が前、左が後)、下は後ろ側から見たもの。発生中期ではニワトリもカメも、筋板が隣接する肋骨の間を背側から腹側に走り(図では上から下へ)、その外側に肩甲骨がある。筋板と肋骨は同じ平面上にある。ただしカメの肋骨はニワトリのものと比べると短く、CR(甲稜)よりも腹側には肋骨が見られない。ニワトリではこの後の発生でも、その形態はほとんど変わらない。発生後期のカメではCR近傍で腹側の体壁が内側へと折れ曲がり、肩甲骨が肋骨の腹側へと入る。しかし、肩甲骨は相変わらず肋骨と筋板でつくられる平面の外側にあり、相対的位置関係はニワトリや発生中期のカメと変わらない。
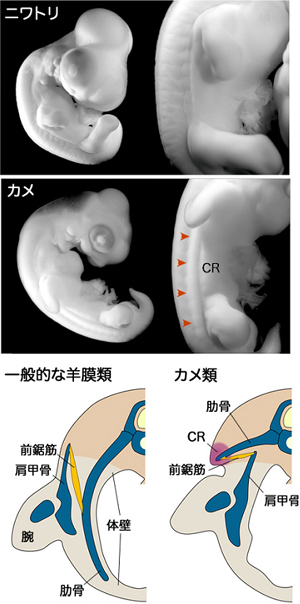
図6 CRと前鋸筋の裏返り
(上)発生中期のニワトリ胚とカメ胚の側面観。カメ胚では発生中期に将来の甲羅の縁となるCR(矢頭)が体側に形成される。
(下)一般的な羊膜類の胚とカメの胚の横断面の模式図。一般的な羊膜類の肩甲骨は、肋骨の外側にある。前鋸筋も肋骨の外側にあり肩甲骨と肋骨を結んでいる。カメの肩甲骨は、背側で横方向に広がった肋骨(=背甲)の内側にあり、前鋸筋も肋骨の内側にある。両者で前鋸筋の肋骨に対する絶対的な位置(肋骨の外側か内側かという意味)は異なるが、肋骨-前鋸筋-肩甲骨のつながる順序(相対的位置関係あるいは形態的相同性)は同じである。このカメにおける前鋸筋の位置の特異性も、体壁の折れ込みによってもたらされる。
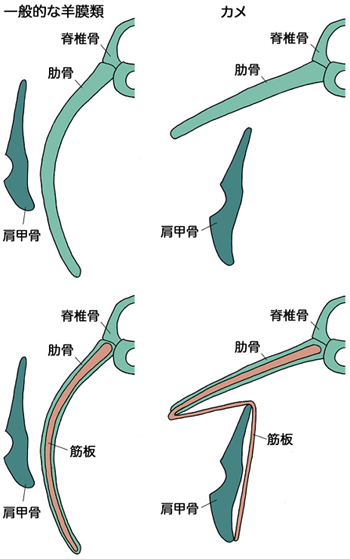
図7 一般的な羊膜類とカメの骨格の位置関係の比較
(上)一般的な羊膜類とカメの肋骨・肩甲骨の横断面。一般的な羊膜類では、肩甲骨が肋骨の外側にあるが、カメでは肋骨の内側にあると考えられ、この位置関係がどのようにして起こったのかが議論の的であった。
(下)上の図に筋板を追加した。本研究で初めてカメにおける筋板の位置を特定した。筋板は一般的な羊膜類でもカメでも肋骨と同じ平面上にあり、またカメでも筋版は肩甲骨の内側を通っている。カメにおける肩甲骨と肋骨の関係については、絶対的な位置関係という意味で、肩甲骨は肋骨の腹側にあるが、筋板を含めた相対的な位置関係という意味では、ほかの羊膜類と同じで、肩甲骨は筋板-肋骨面の外側にある。この特異な位置関係は、筋板の折れ曲がりによってもたらされた見かけ上のものである。
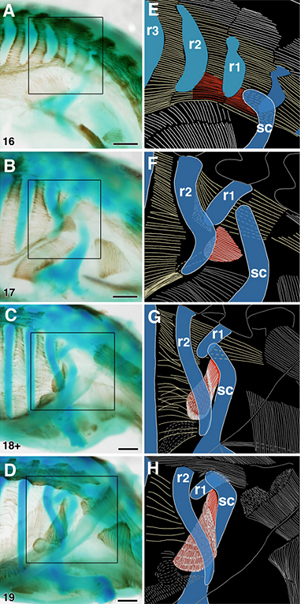
図8 カメの発生に伴う前鋸筋の位置の変化
骨格は染料、筋肉は免疫組織染色したカメ胚の肩の部分の拡大写真(左)と、黒枠の部分を拡大して示した線画(右)。発生中期の段階(A)で、肩甲骨(sc)は一番前の肋骨(r1)の前外側にあり、前鋸筋(赤)が肩甲骨と肋骨との間を前後に走っている(右が前、左が後)。発生が進むにつれて(BからD)、カメの肋骨は前側方に成長し、肩甲骨を後から覆い、同時に前鋸筋はその肋骨へのつながりを維持したまま、裏返りながら、肋骨の内側へと位置を変えていった。略号:r2=第2肋骨、r3=第3肋骨 スケールバー:500μm。

図9 広背筋の発生の比較
カメ、ニワトリ、ハツカネズミにおける広背筋の発生の比較するため、右前足部分でのLbx-1の発現、筋肉および骨格の形態を示した。どの動物でも広背筋の原基(ld)にはLbx-1が発現し(A, E, I)、同じ位置にでき始める(B, F, J)。その後の発生で、ニワトリ、ハツカネズミでは、後ろ(図では左)に向かって筋が伸びてゆくが、カメでは肩甲骨(sc)に沿って、前(図では右)に向かって伸びてゆく。
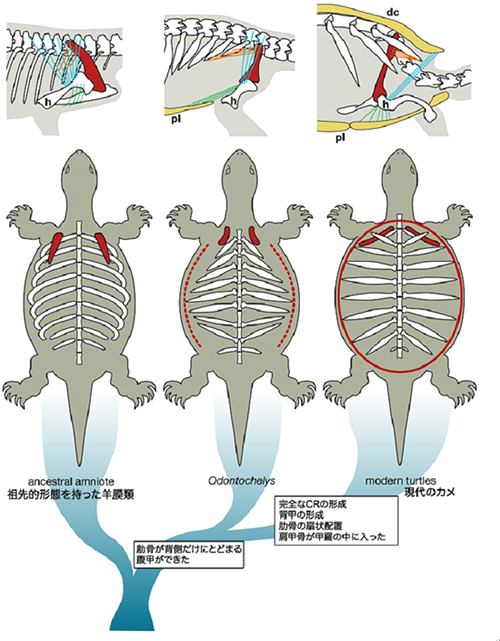
図10 本研究をもとに推定したカメの進化
左から、一般的な羊膜類と同じ筋、骨格の配置を持ったカメの祖先、Odontochelys、現生のカメに至る系統を示す。Odontochelysでは、肩甲骨は肋骨の前外側にあり、肋骨はすでに短く、扇状の広がりを見せずに末端ですぼまった配置をしていた。前鋸筋は肩甲骨と肋骨の間を前後に走っていたと推定、この肩甲骨、前鋸筋、肋骨の形態は、現生のカメの発生中期の胚形態と同じである。腹甲は既にあったので、胸筋は現生のカメと同じように腹甲の内側に付いていたはずである。広背筋の位置は不明。肋骨は扇状の広がりを見せなかったので、CRの機能は現生のカメ胚でのそれと同等の能力はなかったと考えられる。Odontochelysよりも進化した系統でCRの機能が完全となり、発生後期にCRによって肋骨の扇状の配置が起こって背甲ができた。さらに体壁が折れ込んで、前鋸筋、肩甲骨が肋骨の腹側に入り、広背筋は前に伸びて、新たにできた背甲に内側から付着した。