2009年11月12日
独立行政法人 理化学研究所
独立行政法人 科学技術振興機構
視覚経験で抑制性神経回路が「ダイナミック」に変化する現象を初めてキャッチ
-弱視治療で知られる臨界期における眼優位性可塑性を、抑制性細胞の別の可塑性が制御-
ポイント
- 片目を閉じると、特定の抑制性細胞が視覚反応の眼優位性を時間経過で逆転させる
- 興奮性細胞への抑制性入力の役割も、片目を閉じるとダイナミックに変化
- 特定の抑制性細胞の異常で発症する統合失調症、自閉症などの治療にも期待
要旨
独立行政法人理化学研究所(野依良治理事長)と独立行政法人科学技術振興機構(JST、北澤宏一理事長)は、見る聞くなどの感覚的な経験で、脳内の神経回路が機能的に作り変わる「臨界期」と呼ぶ時期に、大脳皮質にある特定の抑制性神経細胞(Fast Spiking細胞※1)が、視覚経験により「ダイナミック」に視覚反応を変化させることを世界で初めて発見しました。理研脳科学総合研究センター(利根川進センター長)神経回路発達研究チームヘンシュ貴雄チームリーダー、杉山(矢崎)陽子客員研究員(JST研究員)、脳回路機能理論研究チーム深井朋樹チームリーダー、姜時友、加藤英之研究員(現・理研BSI-トヨタ連携センター、ユニットリーダー)による研究成果です。
成長初期の脳は、周りの環境や経験による刺激に適応するように、脳内の神経回路を機能的に変化させながら発達していきます。こうした外部からの感覚の入力を基に、神経細胞が機能的に変化することができる時期が「臨界期」です。五感の1つである視覚を例に取ると、脳の高次機能をつかさどる大脳皮質の視覚野(両目からの入力が最初に統合される領域)にある神経細胞は、「見る」という両目からの刺激の入力を基に神経回路を構築していきます。臨界期に片目からの視覚入力を遮断すると、視覚野の80%を占める興奮性神経細胞である錐体細胞※2の視覚反応が、開いている目でより強くなるように神経回路を再構築します。この現象を利用したのが幼児期の弱視治療で、正常な目の方を物理的にアイパッチ(眼帯)でふさぎ、弱視側の目への反応を強制的に強くすることで視力を回復させます。
これまで研究チームは、臨界期の開始には、神経回路内に20%しか存在しない抑制性神経細胞の成熟が必要であることを明らかにしてきました。今回、臨界期に片目を閉じると、10種類以上あるとされる抑制性細胞の中の1つFast Spiking細胞が、錐体細胞とは逆に、2~3日後では閉じた目の方でより強く反応し、14日後には開いている目の方でより強く反応するというダイナミックな視覚反応の逆転現象を、世界で初めて観察しました。また、このFast Spiking細胞の視覚反応の変化が、Fast Spiking細胞から情報を受け取る錐体細胞の視覚反応に影響をもたらし、神経回路全体の可塑性を制御していることを明らかにしました。
本研究は、JST戦略的創造研究推進事業チーム型研究(CREST)の研究領域「脳の機能発達と学習メカニズムの解明」における研究課題「臨界期機構の脳内イメージングによる解析と統合的解明」(研究代表者:ヘンシュ貴雄)の一環として行った成果で、英国の科学雑誌『Nature』(11月12日号)に掲載されます。
背景
外国語や新しいスポーツの習得は、子供の方が早く上手なことから、幼い時に受ける経験は、吸収しやすく定着しやすいことを私たちは経験から知っています。これは、成長過程の特別な時期に、自身の経験や周りの環境からの刺激を受けて、脳内の神経回路が形成、再構築されるためと考えられており、この時期のことを「臨界期」と呼びます。しかし、臨界期という特定の時期に、脳内の神経回路がどのようなメカニズムで集中的に形成されるのか、また、臨界期の開始や終了がどのように決まるのかなどは謎のままです。
研究チームは、この謎を解明するために、哺(ほ)乳類であるマウスの視覚に着目して研究を行っています。大脳の視覚野は、脳の高次機能をつかさどる大脳皮質の中にあり、左右それぞれに存在します。この視覚野は、左右それぞれの目から入った視覚の情報を最初に統合する場所です(図1上)。視覚野を構成する神経細胞は、大きく分けて2種類あり、1つは相手の神経細胞の活動を興奮させる興奮性細胞、もう1つは相手の活動を抑える抑制性細胞です。視覚野の中でこれら神経細胞同士はお互いにつながり合って神経回路を形成しており、神経細胞の80%は興奮性細胞である錐体細胞で、残りの20%は抑制性細胞に分類できることも分かっています。
視覚野両眼領域の神経細胞は、左右どちらの目への光刺激にも反応しますが、通常はどちらか一方の目に強い反応をします。これを「眼優位性」といいます。マウスでは、通常右側の脳の視覚野にある錐体細胞が、左目からの入力に対してより強い視覚反応を示します。逆に、左側の脳の視覚野にある錐体細胞は、右目からの入力に対してより強い視覚反応を示します。つまり、視覚野のある側と反対側の目に優位に反応を示します(図1左下)。しかし、生後の特定の時期である臨界期に左目をふさぐと、錐体細胞の視覚反応はその優位性を右目、つまり開いている目の方が優位になるように神経回路を再構築します(図1右下)。この現象は、弱視の子供の正常な目の方を物理的にアイパッチ(眼帯)でふさぎ、弱視側の目への反応を強くして視力を回復させる弱視治療に応用されています。このように、神経細胞がその反応や形態を状況に合わせて変化させる性質を「可塑性」といい、臨界期は、この可塑性が高い時期ということになります。
研究チームは、これまでの研究から、抑制性細胞の成熟を早めたり遅くしたりすることで、臨界期の開始を人為的に操作することに世界で初めて成功しました(2000年3月7日プレス発表)。さらに、10種類以上ある抑制性細胞の中でもParvalbumin(PV)タンパク質を持つ特定の介在細胞※3(PV陽性細胞※4)の成熟が、臨界期の開始に必要なことも見いだしてきました(2008年8月8日プレス発表)。
しかし一方で、この抑制性細胞自身が、片目を閉じるとどのように視覚反応を変化させるのか、抑制性細胞の「可塑性」については、細胞の数が少ない上に大きさも小さいことから同定が難しく、あまり研究が進んでいませんでした。
研究手法と成果
これまで、視覚反応の可塑性を調べるには、「細胞外記録法」という方法が用いられていました。この方法は、神経細胞の外側から電極を細胞に近づけ、神経細胞が発している電気信号を記録するもので、神経発火※5といった大きな電気信号の記録しか行うことができませんが、比較的簡単に行うことができます。しかし、この方法では、少数で小さい抑制性細胞からの信号を記録することが難しく、また、どの細胞から記録しているのかを同定することは不可能でした。このため、これまでの研究では、神経細胞の80%を占め、大きさの大きい錐体細胞からの記録しか行うことができませんでした。今回研究チームは、細胞内に直接電極を挿入し、1つの細胞の内側から細胞内の膜電位の変化を記録することができる「細胞内記録法」をマウス個体に適用することに成功し、個々の細胞を同定しながら、抑制性細胞の視覚反応や錐体細胞内での抑制性入力の役割を観察しました。
(1)特定の抑制性細胞、Fast Spiking細胞が見せるダイナミックな眼優位性の変化
臨界期の開始に重要な役割をしているPV陽性細胞の多くは、特徴的な非常に速い神経発火が可能な(実験上は40kHz以上)、Fast Spikingの特徴を持つことが知られています。この特徴を持つ細胞をFast Spiking細胞といいます。細胞内記録法を用いると、神経発火の詳細なパターンを観察することができるため、容易にFast Spiking細胞を見分けることができます(図2上)。驚くべきことに、マウスのFast Spiking細胞の眼優位性の可塑性は錐体細胞の示すものとはまったく異なり、両目が開いている状態では優位性がほとんど無く(どちらの目にも同じくらいの強さで反応する)、片目を閉じて2~3日後には、閉じた目の側に優位性が移行することが分かりました。さらに目を閉じ続けていると、14日後には開いている目に優位性が移行しました(図2下)。つまり、錐体細胞は、目を閉じることで開いている側の目へ一方的に優位性が移行するのに対し、抑制性細胞の1つであるFast Spiking細胞は、初めは閉じた側の目に、その次に開いている側の目へとダイナミックにその優位性を逆転させることが初めて分かりました。
(2)Fast Spiking細胞の可塑性の規則と神経回路のシミュレーション
次に、どのようにしてFast Spiking細胞がダイナミックな眼優位性の逆転を引き起こすのかを理解するため、シミュレーションによる実験結果の再現を試みました。Fast Spiking細胞と左右それぞれの目からの入力経路のモデルを構築し、それぞれの目からの入力を与えるシナプス(神経細胞同士が結合し、情報を伝える部位)に、入力信号の到達時刻と、それを受ける細胞の発火時刻との時間差に応じて結合強度が変化するような規則(スパイクタイミング依存可塑性、Spike Timing Dependent Plasticity:STDP)を適用させてシミュレーションを行いました。その結果、細胞内記録法で観察したのと同様に、モデルのFast Spiking細胞が受けるシナプスにおいてもダイナミックに優位性が移行する様子を再現することができました(図3)。
(3)単眼遮蔽による錐体細胞での抑制性入力の役割の変化
大脳皮質の視覚野では、錐体細胞と抑制性細胞が結合し、複雑な神経回路を形成しています。つまり1つの錐体細胞における視覚反応は、興奮性の入力と抑制性の入力が統合することで決定します。そこで、抑制性細胞であるFast Spiking細胞で見つかったダイナミックな眼優位性の逆転が、抑制性細胞から信号を受け取る側の錐体細胞へどのように反映するのかを調べました。つまり、錐体細胞の眼優位性の決定における抑制性入力の役割が、片目を閉じるとどのように変化するのかを観察しました。
具体的には、抑制性の神経伝達物質γ-アミノ酪酸(GABA)※6の情報伝達を阻害する薬剤を充填した電極を用いて、錐体細胞の膜電位を細胞内記録法で測定しました。細胞内記録法で用いる電極の先端は非常に細いため、薬剤はその先端から記録中に少しずつ細胞の中に漏れ出ていきます。つまり、記録を始めた直後では薬剤の影響のない視覚反応を、長時間(30分以上)経過した後では薬剤の影響下(抑制性入力の阻害下)での視覚反応を見ることができます(図4上)。すなわち、この2つを比較すると、同一の細胞内で抑制性入力の役割が分かります。
実験の結果、錐体細胞の視覚反応における抑制性入力の役割が、片目を閉じることにより大きく変化することを明らかにすることができました。錐体細胞内では、通常、抑制性入力が視覚反応の優位性を大きくするように働きます。しかし、片目を閉じて2~3日後には、優位性のある目を逆転させるように働き、この働きは14日後でも弱く残っていました(図4下)。
さらに、錐体細胞における抑制性入力の役割の変化が、視覚野の神経回路におけるどの結合変化に起因しているのかを明らかにするため、数タイプの神経回路モデルを構築し、それぞれシミュレーションを行って、実験結果が再現できる条件を調べました。その結果、細胞内記録法による実験から得た薬剤による視覚反応の変化を再現するためには、左右の眼から抑制性細胞へのシナプス結合の強さの変化が必須であることを明らかにすることができました(図5)。
今後の期待
今回の研究から、視覚野の神経回路を構成する少数派の抑制性細胞のうちFast Spiking細胞が、片目を閉じることにより、閉じた側の目から開いた側の眼へと、ダイナミックに眼優位性を逆転させることを発見しました。神経回路内のFast Spiking細胞の視覚反応の変化が、神経回路全体の働きを制御し、従来知られている眼優位性の可塑性を作り出していると考えられます。
抑制性細胞は多種多様で、それぞれ違った役割があるといわれています。そのため、抑制性細胞の種類を同定しながら詳細に解析していくことで、各種の抑制性細胞の重要な役割が明らかになり、臨界期のメカニズムを理解できると考えています。本研究で注目したFast Spiking細胞は、主にPV陽性細胞であり、統合失調症、自閉症といった精神疾患にはPV陽性細胞に異常のあることが分かっています。特定の抑制性細胞の臨界期における役割、そのメカニズムを解明することで、弱視だけでなく、精神疾患の治療などにも新しい知見をもたらすと期待できます。
発表者
理化学研究所
脳科学総合研究センター 神経回路発達研究チーム
チームリーダー Takao K. Hensch(ヘンシュ貴雄)
Tel: 048-467-9634 / Fax: 048-467-2306
客員研究員 杉山 陽子(すぎやま ようこ)
Tel: 048-467-9673 / Fax: 048-467-2306
お問い合わせ先
脳科学研究推進部 企画課納富 さより(のうどみ さより)
Tel: 048-467-9757 / Fax: 048-462-4914
(JSTの事業に関すること)
独立行政法人科学技術振興機構
イノベーション推進本部 研究領域総合運営部
廣田 勝巳(ひろた かつみ)
Tel: 03-3512-3524 / Fax: 03-3222-2064
報道担当
独立行政法人理化学研究所 広報室 報道担当
Tel:048-467-9272 / Fax:048-462-4715
独立行政法人科学技術振興機構 広報ポータル部
Tel: 03-5214-8404 / Fax: 03-5214-8432
補足説明
- 1.Fast Spiking細胞
電気生理学的に、非常に高い頻度で、しかも減衰することなく発火し続けることができる特徴を持つ細胞のこと。PV(Parvalbumin:パルバルブミン)陽性細胞のほとんどはFast Spiking細胞であるといわれている。 - 2.錐体細胞
大脳皮質の神経細胞のうち、約80%を占める興奮性神経細胞。大脳皮質の神経細胞や脳のほかの部位の神経細胞と互いに結合し、主に脳内の情報のやりとりを担う。この細胞がやりとりする情報の量や質は、介在細胞によって細やかに制御される。 - 3.介在細胞
大脳皮質の神経細胞のうち、約20%を占める抑制性神経細胞。錐体細胞の担う情報伝達を的確に抑制し、大脳皮質内の神経回路(局所神経回路)の活動を制御する。多種多様で細胞の形や機能の違いから、10種類以上の細胞があると考えられている。細胞の種類を見分けるために、PV(Parvalbumin:パルバルブミン)タンパク質など個々の細胞種が持つ固有のタンパク質を検出する方法がよく用いられる。 - 4.PV陽性細胞
大脳皮質の介在細胞のうち、PV(Parvalbumin:パルバルブミン)タンパク質を内在する抑制性神経細胞。介在細胞の約50%を占める。この細胞の形と機能から、錐体細胞の活動を強く抑制すると考えられている。 - 5.神経発火
1つの神経細胞が発する大きく速い、Na+イオンとK-イオンの出入りに依存した膜電位の変化。この信号が結合する別の神経細胞に伝達されていく。 - 6.γ-アミノ酪酸(GABA)
大脳、小脳や海馬の主要抑制性伝達物質。脳内GABA濃度の低下はけいれんを誘発させ、病態との関連が注目されている。
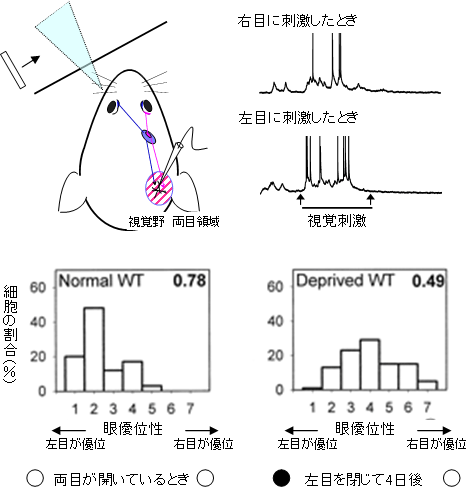
図1 視覚野にある神経細胞の視覚反応と「臨界期」における眼優位性の可塑性
上側:両目で物を見たとき、両目からの視覚情報は大脳皮質の視覚野の両眼領域において初めて統合される(左上 斜線部)。視覚野(両目領域)の細胞が、どちらの目により強い反応をするか(眼優位性)を検出するために、片目ずつに光刺激を与え、細胞が発する電気信号を記録する(右上)
下側:右側にある視覚野の細胞が、左目の光刺激により強く反応した場合(グループ1,2,3)から、右目により強く反応した場合(グループ5,6,7)まで、その優位性に応じて細胞をグループ分けする。両目に均等に反応した場合はグループ4になる。普通のマウスの右側の視覚野にある神経細胞は、左目に優位な反応をする細胞が多いが(左下)が、「臨界期」に左目を閉じると、開いている右目により強く反応する細胞が増える(右下)。
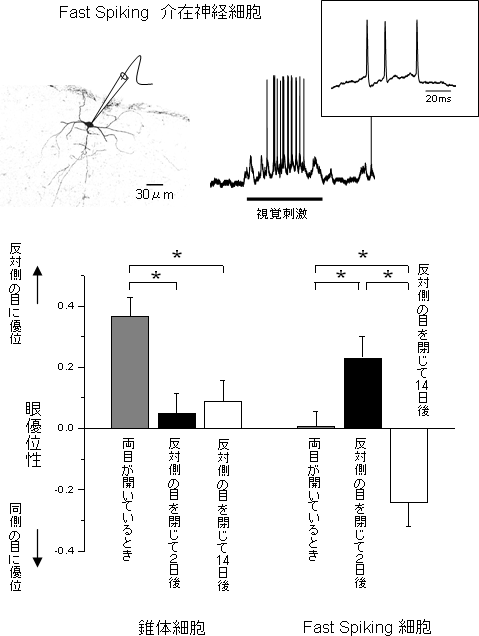
図2 Fast Spiking 細胞の眼優位性の可塑性
上側:大脳視覚野には興奮性の錐体細胞と抑制性細胞がある。抑制性細胞はさらに細かく10種類以上に分類される。その中の1つにFast Spikingという高い頻度で発火する細胞群がある
下側:片目を閉じると、錐体細胞は開いている目(同側の目)の優位性が高くなる方向に眼優位性は移行する(反対側の目を閉じるので図では下向きの方向)が、驚くべきことにFast Spiking細胞は、初め(2日後)は閉じた目(反対側の目)により強い反応をするように優位性が動き、14日経つと、開いている目(同側の目)の優位性が高くなる
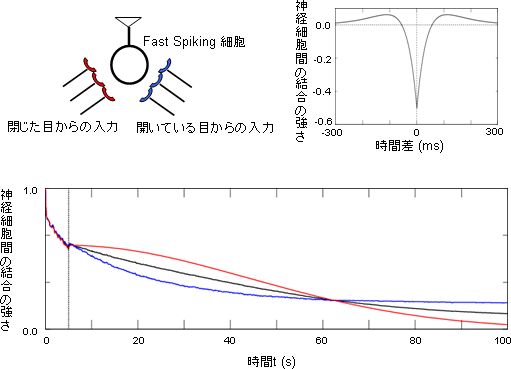
図3 Fast Spiking細胞のダイナミックな眼優位性の可塑性を再現するシミュレーション
上側:抑制性細胞(Fast Spiking細胞)は、閉じた目からも開いたままの目からも複数のシナプス入力を受ける。個々のシナプス入力が与えられた時刻と、それを受けるFast Spiking細胞が発火した時刻との時間差に応じて、対応するシナプスの結合強度が変化する。これをスパイクタイミング依存可塑性(Spike Timing Dependent Plasticity:STDP)という。時間差によってはシナプスが増強することも減弱することもある(正の値が増強、負の値が減弱)。
下側:スパイクタイミング依存可塑性のルールに従いシミュレーションを行って得た、それぞれの眼からの入力の結合強度の時間変化(赤:閉じた眼からの入力強度の時間変化、青:開いている眼からの入力強度の時間変化、黒:両眼の平均)。
t=5秒において片方の眼を閉じると(黒点線)、それ以降はまず閉じた方の眼からの入力に対してより強い視覚応答を示すが(赤>青)、ある時間経過の後には(t>70秒付近)開いている眼からの入力に対してより強い視覚応答を示す(赤<青)。
これは細胞内記録法の結果と一致しており、片目を閉じた後の時間経過とともにそれぞれの眼からの入力の結合の強弱関係が逆転するという現象が、シミュレーションによっても再現できた。
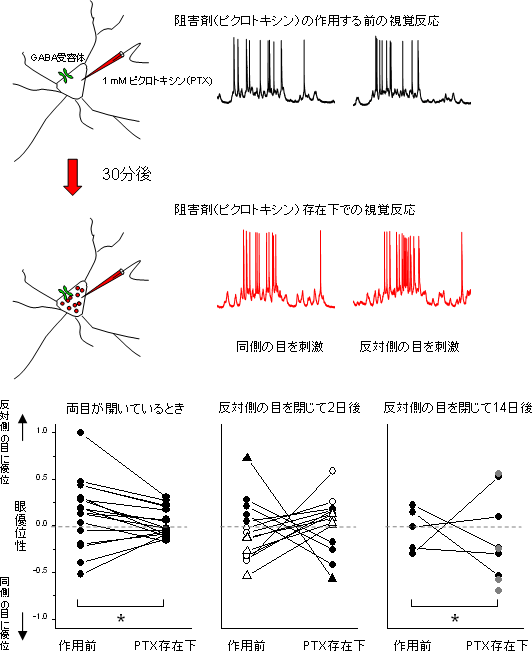
図4 錐体細胞に対する抑制性入力の変化
上側:抑制性の入力を受け取るGABA受容体の阻害剤であるピクロトキシン(PTX)を入れた電極で、錐体細胞の細胞内記録を長時間行い、電極の先端から少しづつ阻害剤を細胞内に充満させる。錐体細胞内に電極が刺入直後と30分以上経った後、つまりPTXの作用前と作用後の視覚反応の眼優位性を比較すると、抑制性入力の眼優位性における役割を知ることができる。
下側:両目が開いてるときは、PTXの作用により抑制性入力が阻害されて優位性が弱くなるが(左)、片目(記録している視覚野と反対側の目)を閉じて2日後にはPTXの作用によって優位性の高い目が逆転し(真ん中)、さらに14日後には、作用前に視覚反応が無かった細胞がPTXの作用により反応するようになる(右、灰色の丸)。このように、錐体細胞の眼優位性おける抑制性入力の役割が、片目を閉じることによって変化していることが分かった。
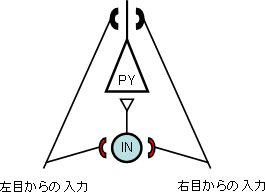
図5 視覚野内の局所回路
視覚野内では錐体細胞(PY)と抑制性細胞(IN)が連絡し合って神経回路を形成しており、両目からそれぞれ入力がある。「臨界期」に片方の目を閉じると、抑制性細胞への入力を担うシナプス結合(赤色の部分)において結合の強さに変化が起こり、その結果として抑制性細胞の視覚反応がダイナミックに変化する。この抑制性細胞への入力部分の変化が、抑制性細胞から情報を受け取る錐体細胞の視覚反応の変化を制御していると考えられる。