2010年12月21日
独立行政法人 理化学研究所
CAPS2が神経栄養因子BDNFの分泌を増強し、脳回路を正常に発達
-CAPS2の分泌増強効果の欠損が、抑制性シナプスの異常を起こし不安を亢進-
ポイント
- CAPS2が持つ神経栄養因子BDNFの分泌増強効果を、イメージング技術で解明
- BDNF分泌の増強が、海馬のGABA作動性の抑制性神経回路発達に重要
- 不慣れな環境下では、CAPS2遺伝子欠損マウスの不安行動が亢進
要旨
独立行政法人理化学研究所(野依良治理事長)は、蛍光イメージング解析により、刺激を受けた神経細胞の分泌顆粒が放出するBDNF(脳由来神経栄養因子)※1の量が、分泌調節因子CAPS2※2の働きによって増強する動態を初めてとらえることに成功しました。これは、理研脳科学総合研究センター(利根川進センター長)分子神経形成研究チームの古市貞一チームリーダー、篠田陽客員研究員らの研究成果です。
BDNFは、神経細胞の生存・分化、神経回路の発達・機能などの調節に必須な分泌性のポリペプチドで、精神神経疾患(うつ病、統合失調症、発達障害、アルツハイマー病など)との関連も示唆される重要な神経栄養因子の1つです。このBDNFは、神経細胞の軸索※3に分布する分泌顆粒「有芯小胞※4」に含まれており、脱分極刺激※5を受けると有芯小胞が細胞膜外に開口してBDNFを分泌(開口放出※6)します。この開口放出には、CAPS2が関与すると考えられていましたが、これまでCAPS2がBDNFの分泌をどのように増強するのか、神経細胞上で促進している様子やその時系列的変化については不明のままでした。
研究チームは、CAPS2遺伝子欠損マウスの海馬※7由来の神経細胞を用いて、BDNFの分泌を蛍光イメージングで解析したところ、CAPS2の存在がBDNF分泌の動的速度を約30%、頻度を約85%、量を約60%増強していることを明らかにしました。また、CAPS2遺伝子欠損マウスでは、海馬のGABA(γアミノ酪酸)※8作動性の抑制性神経回路が脆弱になるとともに、抑制性シナプス発達、抑制性シナプス電流、シナプス可塑性、脳波に異常を示し、さらに、新奇オブジェクトを設置した不慣れな環境下で不安様行動を亢進することも明らかにしました。
今回の研究成果は、CAPS2の分泌増強効果の欠損が、BDNF分泌の減弱を引き起こしてGABA作動性の抑制性神経回路を脆弱にし、全般性不安障害やパニック障害といった不安障害※9を発症するリスクの可能性を示しました。今後、不安障害だけでなく、BDNFとの関連が示唆されるうつ病や統合失調症、自閉症の一種であるレット障害、アルツハイマー病などの精神神経疾患でも、CAPS2の分泌増強効果を利用した臨床への新しい応用・開発が期待できます。
本研究成果の一部は、独立行政法人科学技術振興機構 戦略的創造研究推進事業チーム型研究(CREST)の研究課題「BDNF機能障害仮説に基づいた難治性うつ病の診断・治療法の創出」(研究代表者:小島正己 独立行政法人産業技術総合研究所 研究グループ長)の分担研究によって得られ、米国科学アカデミー紀要『Proceedings of the National Academy of Sciences of the United States of America: PNAS』12月20日の週にオンライン掲載されます。
背景
脳を構成する神経細胞には、小型(直径約50nm)のシナプス小胞※4と大型(直径約80~120nm)の有芯小胞という2つの袋状の構造をした分泌小胞が存在します。有芯小胞には、カテコールアミン(ドーパミンやノルエピネフリンなど)や神経ペプチドが含まれており、この有芯小胞の膜が細胞外に開口して分泌物質を放出します(開口放出)。脳由来神経栄養因子BDNFもこの有芯小胞の開口放出で分泌され、約1,300個のアミノ酸からなる分泌調節因子CAPS2が、この過程に作用すると考えられています(図1)。BDNFは、神経細胞の生存と分化、神経回路の形成、シナプスの可塑性などを調節する上で必須な生理活性を持っており、精神神経疾患との関連も示唆されている重要な神経栄養因子の1つです。これまで研究チームは、CAPS2がBDNFを含む有芯小胞の膜に会合した後、例えば高濃度な塩化カリウム(KCl)処理により有芯小胞が脱分極刺激を受けると、細胞内Ca2+が増加してBDNFの分泌活性が促進すること、CAPS2遺伝子欠損マウスが神経細胞やシナプスの発達異常、社会性行動の欠損や不安様行動の増加を発症すること、さらには自閉症患者の中にCAPS2亜型の発現に異常を持つ人が存在すること、などを報告してきました(2007年3月23日プレス発表)。
しかし、どのようにCAPS2がBDNFの分泌を促進するのか、分泌を制御するメカニズムや細胞における分泌の動態についての詳細は不明でした。また、CAPS2のBDNF分泌増強効果が、細胞生物学的にどのような影響を及ぼすかということもよく分かっていませんでした。研究チームは、発達障害や不安障害に関係する脳回路の正常な発達と働きを理解する上で、これらの問題を解決することが重要であると考えました。
研究手法と成果
(1)神経細胞におけるBDNF分泌のイメージング
神経細胞の内と外のpHの値は約7と中性に保たれていますが、分泌小胞の内部は約5.5と一般的に酸性に偏っており、この性質を利用して有芯小胞内外のBDNFをイメージングすることができます(図2)。具体的には、BDNFとpH感受性蛍光タンパク質(pHluorin)を融合した組み換えタンパク質「BDNF-pHluorin」をマウスの海馬由来の神経細胞に発現させます。このBDNF-pHluorinは、酸性の有芯小胞内にあるときは消光しており、中性である細胞外に放出されるとその放出部位で斑点状の蛍光を発します。高濃度KClで神経細胞を脱分極刺激すると、有芯小胞が開口してBDNF-pHluorinを細胞外に放出するので、刺激前後にわたって、神経細胞の神経突起上で微弱な蛍光の出現と変化を観察することができます。この蛍光斑点の経時的な動態を高感度なカメラで記録しました。
実験には、CAPS2遺伝子欠損マウスの海馬由来初代培養神経細胞(CAPS2-)を使用しました。この細胞にCAPS2遺伝子を外部から導入し(CAPS2+)、CAPS2-細胞と比べました(図3)。CAPS2+細胞とCAPS2-細胞にBDNF-pHluorinを発現させて、脱分極刺激によって発生する蛍光がピークになるまでの時間、その動的速度(時定数)と頻度、そして蛍光斑点の一定時間当たりの数とシグナル強度(分泌量に相当)などについて比較しました。その結果、CAPS2+細胞の場合は、一定時間当たりのBDNFの動的速度(時定数)を約30%加速(図3B)、分泌頻度を約85%増加(図3C)、1つの蛍光斑点あたりのシグナル強度(分泌量に相当)を約60%増大(図3D)することが分かりました。
(2)海馬GABA抑制系の細胞生物学的な意義
CAPS2遺伝子欠損マウスの海馬由来の神経細胞を解析したところ、興奮性よりも抑制性の神経細胞の異常が顕著であることが分かりました。具体的には、正常マウスと比べてCAPS2遺伝子欠損マウスの海馬CA1領域では、GABA作動性の抑制性神経細胞の数が生後28日で約20%減少し(図4A)、GABA抑制性シナプスの数も約20%減少することが分かりました。また、電子顕微鏡による微細組織形態の解析により、抑制性シナプスにおけるシナプス小胞の数が生後56日で約35%減少し、シナプス小胞分布領域も約30%減少(図4B)することも分かりました。さらに、海馬の急性スライス組織標本を用いた電気生理学的な解析により、抑制性シナプス後電流※10の頻度が約65%、振幅が約20%減少すること(図4C)、海馬CA3領域からCA1領域へ投射されるシナプス間の可塑性が低下すること、この可塑性の低下はGABA受容体阻害剤を添加すると正常マウスと同じ増強レベルになること、なども見いだしました。生きたマウスの海馬からの脳波も観測したところ、CAPS2遺伝子欠損マウスの場合、覚醒中のシータ波というGABA抑制系と関係する脳波の頻度が減少することも明らかになりました。
これらの結果は、CAPS2の分泌増強効果の欠損によってGABA作動性の抑制性神経回路が脆弱になることを示唆しています。抑制性神経細胞の生存と分化には、BDNFが重要であることから、CAPS2はBDNF分泌を増強することによって、抑制性神経回路の正常な発達と機能発揮を促している、という細胞生物学的な意義が分かりました。
(3)マウス行動解析
正常マウスとCAPS2遺伝子欠損マウスの不安様行動評価、学習・記憶評価、うつ様行動評価といった行動表現型について解析を行いました。その結果、CAPS2遺伝子欠損マウスは、高所の十字アーム(高架式十字迷路テスト)では塀の無いアームには入らない、不慣れな環境下での給餌(新奇性抑制摂食テスト)では、空腹であっても摂食までの潜時が長い、8つあるアームでのアーム選択回数(8方向放射状迷路テスト)が少ない、Y字迷路でのアーム選択回数(Y字迷路テスト)が少ないといった不安様行動の増加を示しました。また、うつ様行動のテストでは、水が嫌いなマウスを水槽中に入れた時(強制水泳テスト)に、逃避行動を諦めて無動状態になる時間が長いという兆候を示しました。しかし、ほかのテストでは差が無かったことから、うつ様ではないものの、その傾向はあることが示唆できました。
今回、海馬神経細胞の蛍光イメージング解析を駆使した結果、脱分極刺激を受けた神経細胞で起きる有芯小胞の開口放出によるBDNF分泌の動的速度、頻度、量は、CAPS2によって増強されるという動態を初めて明らかにすることができました。また、組織形態学的および電気生理学的解析の結果、CAPS2の分泌増強効果の欠損がGABA作動性の抑制性神経回路を脆弱にし、海馬の抑制性シナプスの発達、抑制性シナプス電流、シナプス可塑性、脳波などでも異常を示すことが判明しました。また、CAPS2遺伝子欠損マウスは、新奇環境下などで不安様行動の亢進を示すことも明らかにすることができました。これらの結果は、CAPS2の分泌増強効果の欠損によりBDNFからGABAにつながる神経回路が脆弱になると、不安障害の発症リスクが高まる可能性を示しています(図5)。
今後の期待
この研究成果を糸口にして、脳の発達と働きにおける神経調節因子の分泌制御の基礎研究分野がさらに発展することが期待できます。不安障害の薬物療法では、GABA受容体作動薬が抗不安薬として精神安定作用を示します。また、BDNF分泌量の増減が精神疾患(うつ病、統合失調症、発達障害など)や神経変性疾患(アルツハイマー病やハンチントン病など)に関連するという報告が数多くあります。今後、CAPS2の分泌増強効果を利用した臨床への新しい応用・開発が、発達障害や不安障害のみならず精神神経疾患に対しても期待できます。
発表者
理化学研究所
脳科学総合研究センター 分子神経形成研究チーム
チームリーダー 古市 貞一(ふるいち ていいち)
Tel: 048-467-5906 / Fax: 048-467-6079
お問い合わせ先
脳科学研究推進部 企画課Tel: 048-467-9757 / Fax: 048-462-4914
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.BDNF(脳由来神経栄養因子)
BDNF(ビーディーエヌエフ)は神経栄養因子の1つで、119個のアミノ酸からなる分泌性のポリペプチドである。哺乳類の神経栄養因子にはBDNFのほかにNGF、NT-3、NT-4/5がある。分泌された神経栄養因子はターゲットとなる細胞膜上の特異的な受容体型チロシンキナーゼに作用して、細胞内シグナル伝達を活性化し、神経細胞の生存・分化、神経突起形成などの調節作用を持つ。BDNFはシナプス可塑性、神経回路の発達、活動依存的な遺伝子発現の誘導などにも関与し、BDNF欠損マウスは生後数週間以内で死亡する。また、うつ病、統合失調症、発達障害(レット障害など)、パニック障害、アルツハイマー病、ハンチントン病などでBDNF量の変化が報告されており、精神神経におけるBDNFの重要性が示唆される。しかし、BDNFの分泌制御メカニズムや分泌動態については不明な点が多い。 - 2.CAPS2
Ca2+-dependent activator protein for secretion 2の略称。遺伝子名はCAPS2またはCadps2と表記される。タンパク質ファミリーには、ほかにCAPS1がある。当初CAPS(キャップス)は、神経伝達物質のカテコールアミン(ドーパミン、ノルエピネフリンなど)や神経ペプチドなどを含有する有芯小胞の開口放出にかかわる分泌調節因子として同定された。有芯小胞の開口放出のメカニズムは、シナプス小胞の開口放出に類似していると考えられているが、関与する分子についてはよく分かっていない。研究チームはこれまでに、CAPS2が神経栄養因子であるBDNFとNT-3の分泌を促進することを世界で初めて示した。 CAPS1遺伝子欠損マウスは生後間もなく致死となり、 CAPS2遺伝子欠損マウスは神経細胞の発達異常により社会性行動の欠損と不安行動の増加を発症する。研究チームは、自閉症患者で突起輸送が欠損したCAPS2亜型の異常発現を発見しており、患者ゲノムでCAPS2のコピー数多型も見いだされている。このように、CAPS2は神経調節に非常に重要であり、さまざまな精神神経疾患に関連する分泌因子の分泌作用に関係しているが、その機能的および細胞生物学的な役割の詳細には不明な点が多い。 - 3.軸索
神経細胞体から伸びる神経突起の1つで、出力線維である。軸索終末には神経伝達物質を含有するシナプス小胞やペプチド、カテコールアミンなどを含む分泌顆粒があり、標的となる神経細胞の樹状突起や細胞体とシナプス結合して信号を送る。 - 4.有芯小胞、シナプス小胞
神経細胞の伝達物質の分泌に関与する分泌小胞には、大別してシナプス小胞と有芯小胞の2つがある。シナプス小胞は小型の分泌小胞(直径約50nm)で、低分子の神経伝達物質(アセチルコリン、グルタミン酸、GABA、ドーパミン、ノルエピネフリン、セロトニンなど)の開口放出に関係する。一方、有芯小胞は電子顕微鏡下で芯があるように見える大型の分泌小胞(直径80~120nm)で、神経ペプチドやカテコールアミン系の伝達物質(ドーパミン、ノルエピネフリンなど)などの開口放出に関係する。BDNFは有芯小胞に含まれて輸送・分泌される。 - 5.脱分極刺激
神経細胞は、特異的なポンプなどの働きにより細胞の内外でプラスとマイナスのイオン濃度に差があり(例えば、ナトリウムイオン:Na+は細胞の外が高く内が低い。逆にカリウムイオン:K+は外が低く内が高い。塩素イオン:Cl-は外が高く内が低い。)、これによって電気化学的な勾配が生じて(膜電位)、静止状態で細胞膜の内側が-60~-70mVとマイナス側に分極した電位で平衡に保たれている。シナプス伝達で、グルタミン酸などの興奮性神経伝達物質が伝わると、特異的なイオンチャネル型受容体が活性化され、チャネルが開いてプラスのNa+が細胞の外から内に透過し、その結果分極がプラス側にシフトする。強いシナプス伝達ではより大量のNa+が流入するため、膜電位は0mVを超えて、さらに+40mV付近に向かってシフトする。これを脱分極といい、神経細胞膜が興奮している状態である。脱分極によって電位依存性のカルシウムイオン(Ca2+)チャネルが活性化されCa2+が細胞内に流入すると、Ca2+依存的なシナプス小胞の開口放出の誘導やさまざまなシグナル伝達の引き金となる。神経細胞を電気刺激したり、細胞外を高濃度の塩化カリウム(KCl)溶液に変えることでも人為的に脱分極を誘導できる。一方、抑制性の神経伝達物質であるGABAは、Cl-を流入させ、膜電位をよりマイナス側にシフトさせる。これを過分極という。 - 6.開口放出
神経伝達物質を含むシナプス小胞や、カテコールアミンや神経ペプチドを含む有芯小胞が、細胞膜と特異的に膜融合することにより小胞内腔と細胞外がつながり、含有されていた分泌物質が細胞外へと放出される現象をいう。分泌部位に輸送されてきた小胞は、細胞膜の特異的な部位(シナプス小胞の場合はアクティブゾーン)に結合して(ドッキング)、準備完了になり(プライミング)、刺激が与えられると小胞膜と細胞膜が融合する(フュージョン) (図1参照)。シナプス小胞の開口放出の過程には、小胞膜と細胞膜に存在するSNARE(スネア)と呼ばれる特異的なタンパク質複合体など多くの因子が関係する。有芯小胞の開口放出の機構にもシナプス小胞と同じようなメカニズムが働くと予想されているが詳細は分かっていない。CAPS2は有芯小胞の分泌経路に関与しており、開口放出のプロセスで重要なはたらきをしているとも示唆されている。 - 7.海馬
大脳側頭葉の内下部にあり、両側を合わせた形がギリシャ神話の海神がまたがる海馬に似ていることからこの名称が付いた。両側を破壊すると記憶障害が起きることから、記憶や空間学習に関与すると考えられている。 - 8.GABA(γアミノ酪酸)
GABA(ギャバ)は、脳に存在する神経細胞間のシナプス伝達において、プレシナプス(神経伝達物質を送り出す側)とポストシナプス(神経伝達物質を受け取る側)の間の化学シナプスではたらく抑制性の神経伝達物質の1つである(興奮性の神経伝達物質はグルタミン酸)。GABAは、シナプス小胞の開口放出でプレシナプスから放出され、ポストシナプス膜上のイオンチャネル型受容体(GABA-A受容体)に作用して負電荷の塩素イオン(Cl--)イオンの透過を誘導し、神経細胞の膜電位をよりマイナス側に低下させて(過分極)、神経細胞の興奮を抑制する。また、細胞内シグナル伝達を活性化する代謝型受容体(GABA-B受容体)にも作用し、カリウムイオン(K+)チャネルを介して同様に神経細胞の興奮を抑制する。GABA抑制系は、脳発達期における神経回路形成でも重要な役割を果たすことが知られている。 - 9.不安障害
全般性不安障害、パニック障害、恐怖症、強迫性障害、外傷後ストレス障害、急性ストレス障害、物質誘発性不安障害など不安を主症状とする疾患群の総称で、特徴的な不安症状を呈するもの、トラウマなどが原因のもの、病気や物質によるものなどさまざまである。わが国では、なんらかの不安障害の生涯有病率が約9%と高く、米国でも年々増加傾向にあり、大規模調査によると10人に3人以上が経験する。うつ病などでは不安障害を併存する場合がある。原因は十分には解明されておらず、心理的、社会的な要因、脳機能に関係する生物学的要因が考えられている。薬物療法では抗うつ薬と抗不安薬を処方しており、抗うつ薬には選択的セロトニン再取り込み阻害薬SSRIなど、抗不安薬には精神安定作用を持つGABA受容体作動薬のベンゾジアゼピン誘導体などがある。 - 10.抑制性シナプス後電流
抑制性シナプスにおいて、プレシナプス(シナプス前)から分泌されるGABAをポストシナプス(シナプス後)にあるGABA受容体が受け取ると、受容体のチャネルが開いて負電荷の塩素イオン(Cl-)が細胞内に流入することで発生する電流。神経細胞の膜電位をよりマイナス側に低下させ(過分極)、神経細胞の興奮を抑制する。
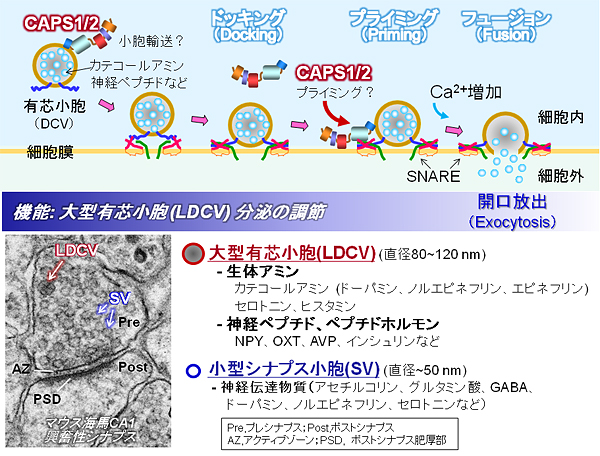
図1 CAPS2は大型有芯小胞の開口放出を調節する
CAPS2とCAPS1は、約1,300アミノ酸からなるタンパク質で、大型の有芯小胞の活動依存的な分泌を調節することが知られている。主にカテコールアミンや神経ペプチドなどが有芯小胞によって分泌されるが、CAPS2はBDNFを含む有芯小胞の分泌を促進することが分かっている。シナプス小胞の開口放出には、SNAREタンパク質複合体をはじめとする多くの調節因子がCa2+依存的な開口放出を調節することが示されているが、有芯小胞の開口放出メカニズムの詳細は不明である(開口放出のプライミングの促進作用や、有芯小胞の輸送での作用などが示唆されている)。また、細胞刺激後に誘導される有芯小胞からの分泌の動態(時系列的変化など)についても不明な点が多く残されている。
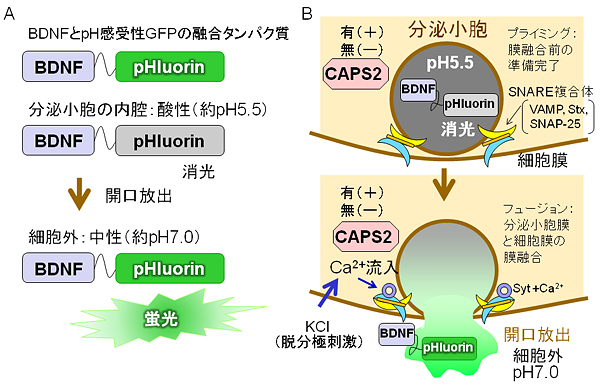
図2 CAPS2によるBDNF分泌の増強のイメージング法
A. BDNFを蛍光イメージングするために、BDNFとpH感受性GFP(pHluorin)の融合タンパク質「BDNF-pHluorin」を作製する。pHluorinは中性条件下である細胞内外では蛍光を示すが、酸性条件下である分泌小胞の内腔では消光する。
B. 分泌小胞に含まれるBDNF-pHluorinは消光の状態で、開口放出で細胞外に分泌された時に蛍光を発する。この系を用いて、CAPS2が発現する細胞としない細胞で、高濃度KCl処理刺激後に神経突起上で変化するBDNF-pHluorinの蛍光を観察する。高濃度のKCl処理は、神経細胞の脱分極を誘導し、電位依存性カルシウムチャネルを活性化してCa2+流入を引き起こす。このCa2+増加が引き金となり、Ca2+と結合したシナプトタグミン(Syt)が準備完了(プライミング)状態のSNAREタンパク質に作用することで、分泌小胞膜と細胞膜が融合(フュージョン)して、開口放出すると考えられている。
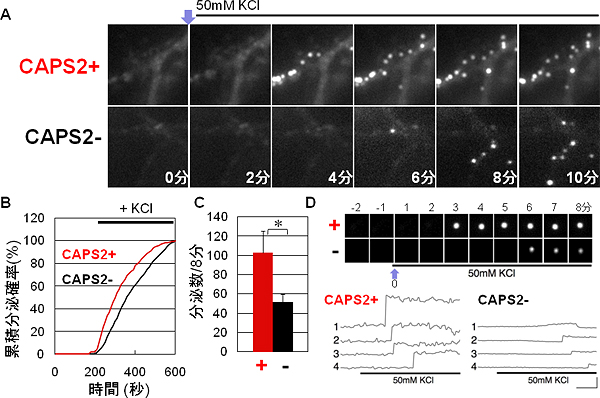
図3 CAPS2によるBDNF分泌動態の増強効果
A)記録開始2分後に、CAPS2遺伝子欠損マウスの海馬由来の神経細胞にKCl刺激を与え、神経突起上でのBDNF-pHluorinの分泌蛍光斑点の動態(2分毎の時系列的変化)を、CAPS2の有無(CAPS2+, CAPS2-)の蛍光イメージングで比較
B)KCl刺激前後の分泌蛍光斑点の累積確率の比較
C)KCl刺激後8分間での分泌蛍光斑点の総数の比較
D)1つの分泌蛍光ごとの動態(1分毎の時系列的変化)と蛍光シグナル強度の比較
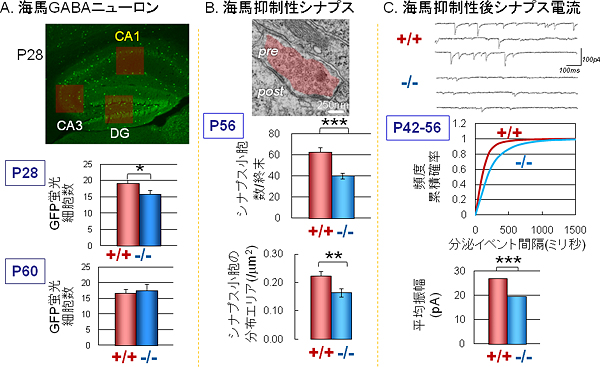
図4 CAPS2のBNDF分泌増強効果の欠失による海馬GABA抑制性神経回路の発達と機能への影響
A. GABA作動性の抑制性神経細胞で蛍光タンパク質(GFP)を特異的に発現する遺伝子改変マウスと交配することによって、蛍光顕微鏡下でこの細胞を観察できる。CAPS2遺伝子欠損マウスの海馬CA1領域における抑制性神経細胞の数を正常マウスと比較すると、生後28日(P28)では減少がみられ、生後60日(P60)ではほぼ同じになる。
B. CAPS2遺伝子欠損マウスの海馬CA1領域における抑制性シナプスを電子顕微鏡観察した。CAPS2遺伝子欠損マウスでは生後56日(P56)でシナプス小胞の数や分布領域の減少がみられる。細胞数はP60で正常と同じ(A)になっても、電顕レベルでは発達異常が残っていることが分かる。
C. CAPS2遺伝子欠損マウスの海馬CA1領域における抑制性シナプス後電流を正常マウスと比較した。CAPS2遺伝子欠損マウスでは頻度と振幅ともに生後42~56日(P42~56)で減少が見られる。従って、成熟した回路の機能レベルでもGABA抑制性シナプスの発達異常は残っていることが分かる。
図5 CAPS2のBDNF分泌増強によるGABA抑制性神経回路の発達への効果と増強作用の欠失が及ぼす精神疾患への影響
正常では、CAPS2はBDNF分泌を増強し、正しいGABA作動性の抑制性神経回路が発達する。CAPS2の異常の場合は、BDNFの分泌増強が欠損し、GABA作動性の抑制性神経回路の発達と機能が損なわれ、その結果、不安障害の発症リスクが高まる可能性が示唆される。CAPS2の異常と自閉症の関連が示唆されることから、この経路はうつ病や発達障害など他の精神疾患の発症リスクにも関係すると考えられる。