2011年3月11日
独立行政法人 理化学研究所
独立行政法人 産業技術総合研究所
植物細胞の脱分化を促進するスイッチ因子を発見
-組織培養の高効率化で、植物の増産や有用物質生産などの応用に期待-
ポイント
- WIND1は、植物の脱分化細胞(カルス)形成を促進する核内スイッチタンパク質
- WIND1遺伝子は、傷口で発現が増大し、植物ホルモンへの応答性を高める
- WIND1遺伝子のオン/オフで、カルスの誘導と根や茎葉への再分化が制御可能に
要旨
独立行政法人理化学研究所(野依良治理事長)と独立行政法人産業技術総合研究所(野間口有理事長)は、傷ストレスを受けた植物がカルス※1(脱分化※2した植物細胞の塊)を形成する際に働くスイッチタンパク質「WIND1」を明らかにしました。これは、理研植物科学研究センター(篠崎一雄センター長)細胞機能研究ユニットの杉本慶子ユニットリーダーと岩瀬哲基礎科学特別研究員、産総研生物プロセス研究部門(鎌形洋一研究部門長)遺伝子転写制御研究グループの高木優研究グループ長らを中心とした共同研究グループの成果です。
農業や園芸分野では、苗のクローン増殖として挿し木法が広く活用されているように、植物の高い器官再生能力は古くから知られています。これは、傷ストレスを受けた植物が傷口の細胞を脱分化させる能力、すなわち細胞の分化※2の度合いを下げて分裂を促進させ、新しく器官を作る準備をする能力に優れているためですが、この分子メカニズムは不明でした。
研究グループは、シロイヌナズナ※3の脱分化した細胞で発現が促進しているWIND1という転写因子※4の遺伝子を同定し、これをシロイヌナズナ植物体で過剰に発現させると、植物体の各器官でカルスが形成されることを発見しました。またWIND1遺伝子は、傷ストレスによって傷口で発現が増大し、植物ホルモン※5の1つであるサイトカイニン※5への応答を高める働きがあることを明らかにしました。またWIND1遺伝子の発現を人為的にオン/オフする実験では、WIND1遺伝子の発現をオンにして一度脱分化させた細胞でこの発現をオフにすると、茎葉や根がカルスから再生(再分化※2)してくる現象を見いだしました。さらに、転写因子WIND1の機能を抑えると傷口での脱分化が抑えられることから、WIND1が植物体の傷口で起こる脱分化を促進するスイッチ機能を有していることが分かりました。
今回の研究は、植物が傷ストレスに応じて脱分化を促進する、という古くから知られていた現象を分子レベルで明らかにした世界初の成果で、効率の良い組織培養※6による植物の増産や有用物質生産などの応用に貢献することが期待されます。
本研究成果は、米国の科学雑誌『Current Biology』3月22日号に掲載されるのに先立ち、オンライン版(3月10日付:日本時間3月11日)に掲載されます。
背景
体に傷ストレスを受けた多細胞生物では、新しく細胞を増殖させて素早く傷口をふさぎ、元通りの組織・器官や新しい別の組織・器官を再生するという現象がよく観察されます。新しい別の組織・器官を再生する時、傷口では、一度分化した体細胞がより低い分化状態になり、細胞分裂能と多分化能(細胞がさまざまな種類の細胞に分化できる能力)を再獲得する「脱分化」というプロセスを経ることが知られています。この細胞の脱分化現象は、植物や一部の動物でも古くから知られていましたが、どのようにして引き起こされるのか、その分子メカニズムは現在でも不明な点が多く、生命科学全体の大きな研究課題の1つとなっています。また細胞の脱分化は、再生医療の基盤となる生命現象であるため、iPS細胞に寄せられる社会的な期待が示すように、現在この分子機構の解明と応用の進展が、国内外で非常に高い注目を集めています。
特に植物は高い再生能力を有していることが古くから知られ、苗の増殖に挿し木法が用いられるなど、広く農業や園芸分野で利用されてきました。さらに、植物体から切り取った組織片の脱分化と再分化には、植物ホルモンであるオーキシン※5とサイトカイニンの濃度比が重要であることが20世紀半ばに明らかとなり、試験管内でのカルス培養法と根や茎葉などの器官への再分化法が確立されてきました。また、タバコの葉の1個の分化した細胞(葉肉細胞)からカルスを経て個体を再生させた歴史的研究から、植物細胞には分化全能性※7があることが明らかとなっています。こうして確立された植物細胞の脱分化・再分化を制御する方法は、新品種作製や有用物質生産などの応用科学に用いられ、種苗産業や花卉(き)・園芸産業などの基盤技術として広く活用されています。しかし、植物細胞の分化全能性そのもの、特に脱分化現象に対する分子レベルの理解は立ち後れていました。
研究グループは今回、植物の脱分化した細胞の遺伝子発現に着目し、この謎に挑みました。
研究手法と成果
(1)植物の脱分化細胞で発現が促進している遺伝子のスクリーニング
研究グループは先行研究として、モデル植物であるシロイヌナズナを用い、脱分化した細胞の中で発現が促進している遺伝子を探索しました。具体的には、植物体と3種類のカルス細胞株に対してDNAマイクロアレイ法による網羅的遺伝子発現解析を行い、分化した植物細胞に比べて3種類のカルス細胞株で共通に発現が促進している遺伝子を選抜しました。その中で、植物細胞でのストレス応答や形態形成に関わる因子が含まれることがよく知られているAP2/ERF転写因子ファミリーに属するRAP2.4遺伝子に注目しました。RAP2.4という名前は、その分子の機能を反映する名前ではなかったため、今回の研究成果に基づいて、この転写因子の遺伝子をWound INduced Dedifferentiation (WIND) 1と名付けました(図1)。
(2)WIND1遺伝子の発現は傷ストレスによって傷口で強く誘導される
遺伝子の発現レベルの変化量を測定するリアルタイム定量PCR法と、遺伝子のプロモーターレポーター解析※8を用いて、WIND1遺伝子が植物体でどのような時に強く発現するかを観察する実験を行いました。その結果、分化した植物体の組織に傷をつけると、数時間でWIND1遺伝子の発現が促進し、WIND1遺伝子の転写産物であるRNAが蓄積することが分かりました。また、遺伝子のプロモーターレポーター解析から、この遺伝子は傷口に近い細胞とそこから生じるカルスで特異的に発現が促進することを明らかにしました(図2)。この結果は、WIND1タンパク質が植物の傷ストレスからのカルス形成機構に何らかの役割を持っている可能性を示すものです。
(3)WIND1遺伝子を過剰発現させるとカルスが誘導
植物体内で過剰に遺伝子発現を誘導するプロモーター(カリフラワーモザイクウィルス35Sプロモーター)を用いて、WIND1遺伝子の機能を調べる実験を行いました。35SプロモーターでWIND1遺伝子を強制的に発現させたシロイヌナズナ植物体では、その発現量の大きさに相関して植物体の成長が阻害され、特に発現量の大きいものは、植物体表面のいたるところにカルス(35S:WIND1カルス)を形成しました(図3)。DNAマイクロアレイ法による解析から、この35S:WIND1カルスでの遺伝子発現のパターンは、通常法で誘導したカルスととてもよく似ていることが分かりました。さらに面白いことに、カルスは通常オーキシンやサイトカイニンなどの植物ホルモンを含む培地で継代培養(一定期間毎に細胞を新しい培地に移して増殖させながら細胞株を維持する培養)しますが、35S:WIND1カルスは、植物ホルモン無添加の培地でも継代培養が可能で、誘導から3年以上経った2011年3月現在も良好に増殖を続けています。この結果は、WIND1タンパク質がカルスの誘導だけでなく、カルス状態の維持にも関わり、同時に、植物ホルモンへの応答に影響を与えている可能性を示すものです。
(4)WIND1遺伝子の発現をオフにするとカルスが再分化
培地へ薬剤を添加して遺伝子発現のタイミングを制御することができる遺伝子発現誘導系を用いて、WIND1遺伝子の機能についてさらに解析を進めました。その結果、WIND1遺伝子の発現誘導系を導入した植物(WIND1発現誘導系植物)でも、WIND1遺伝子の発現で植物体や組織片でカルス形成が促進しました。興味深いことに、このカルスは、誘導剤を含む培地で6カ月継代培養した後に誘導剤を除きWIND1遺伝子の発現誘導を抑えると、低頻度ながら茎葉や根に再分化したことから(図4)、WIND1が誘導したカルスが多分化能を獲得していることが分かりました。さらに、WIND1発現誘導系植物の芽生えを共焦点レーザー顕微鏡で詳細に調べた結果、カルス細胞は胚軸や子葉の表皮細胞からも出てきていることが分かりました。胚軸や子葉の表皮細胞は、種子の中で植物の胚が形成される時にすでに分化し、細胞の特異的な機能を獲得していることが知られています。これらの結果から、WIND1タンパク質が植物細胞の脱分化を促進する因子であることが判明しました。
(5)WIND1タンパク質は傷ストレスで誘導される脱分化を制御する
植物の転写活性化因子※4の機能を転写抑制因子※4に変換し、ドミナントネガティブ※9な表現型を持つ植物体を作り出すCRES-T法※10を用いて、傷害部位におけるWIND1遺伝子の機能を調べました。具体的には、WIND1遺伝子とSRDXと呼ばれる転写抑制ドメインの遺伝子配列をつなげた分子(WIND1-SRDX)を作り、これをWIND1遺伝子自身のプロモーター領域下で発現するようにして植物体に導入しました。これにより、WIND1遺伝子が発現する部位で、機能重複する内在性の他の転写因子の働きも抑えながら、WIND1タンパク質そのものの機能を抑制することが可能になります。この分子を発現する植物体(WIND1機能抑制植物)を用いて傷口でのカルス形成能を評価しました。その結果、野生型の植物体に比べてWIND1機能抑制植物体では傷口からのカルス形成が抑制されていました。培地に植物ホルモン(オーキシンとサイトカイニン)を添加し、カルス形成をより促進させる実験でもカルス形成は抑制されました。これらの結果から、WIND1タンパク質は傷ストレスによって誘導され、傷口で脱分化を促進する分子であることが明らかとなりました。
(6)WIND1タンパク質は植物ホルモンの一種サイトカイニンへの応答性を高める
添加する植物ホルモン(オーキシン及びサイトカイニン)の濃度比を変えた培地で植物組織片を培養し、WIND1タンパク質の植物ホルモン応答への影響を調べました。その結果、WIND1機能抑制植物はサイトカイニン応答の指標である茎葉への再分化を抑えるのに対し、オーキシン応答の指標である根への再分化は抑えませんでした。実際、マーカーを使ってサイトカイニン応答を可視化したところ、傷ストレスを受けた野生型の植物体では、傷害部位で約24時間後からサイトカイニン応答が見られ、48時間後には強いサイトカイニンへの応答活性を観察したのに対し、WIND1機能抑制植物では、48時間以降もサイトカイニンへの応答活性を観察できませんでした。また、サイトカイニン応答系が弱まったシロイヌナズナの遺伝子変異株でWIND1遺伝子を強制的に発現させても、カルス形成の頻度が下がることを確認しました。これらの結果から、WIND1タンパク質は植物ホルモンの一種サイトカイニンへの応答性を高めることで、脱分化を促進することが分かりました(図5)。
本研究では、WIND1がカルス化を誘導する転写因子であることを産総研が発見し、その詳細な機能について理研が解析しました。なお、成果の一部は、独立行政法人新エネルギー・産業技術総合開発機構(NEDO)の「生物機能活用型循環産業システム創造プログラム・省エネルギー技術開発プログラム/植物機能を活用した高度モノづくり基盤技術開発/植物の物質生産プロセス制御基盤技術開発(植物の統括的な遺伝子発現制御機能の解析)」(平成21年度終了)の支援によって得られたものです。
今後の期待
本研究は、植物が傷ストレスに応じて細胞を脱分化させる制御機構の一端を分子レベルで解明し、WIND1タンパク質がそのスイッチとして働く転写因子の1つであることを明らかにしました。具体的には、傷口で発現し機能するWIND1タンパク質が、植物ホルモンの一種であるサイトカイニンへの応答を高め、細胞の脱分化を促進することを突き止めました。植物の傷口でサイトカイニン応答が高められるという可能性を示唆する報告は1986年にありましたが、今回の結果はその予測を分子レベルで明らかにする結果となりました。
研究グループではすでに、WIND1タンパク質の詳細な発現制御機構の解明やWIND1タンパク質が発現を制御する遺伝子の同定という新たな課題に取り組んでおり、植物の複雑な分化全能性発揮の全体像をより明確にすることを目指しています。
さらに、今回の研究成果を基に、種苗産業や花卉・園芸産業、植物を用いた医薬品原料生産産業などの植物系産業で、分子スイッチを利用した効率的な脱分化・再分化技術(分子組織培養)の確立を目指しています。WIND1タンパク質の機能をオンにすることで、通常法では脱分化細胞の誘導と維持が困難な植物種や組織で細胞株の樹立が容易になり、それらの細胞株で細胞の大量増殖をした後に再分化を促進(WIND1タンパク質の機能をオフ)することで、クローン増殖や、有用物質生産が可能になると考えられます。このような技術により、優良品種の大量栽培、飛躍的な生産性向上に向けた分子育種、有用物質生産のための新形質の付与など、多様な植物品種の効率的な育成が可能になると考えています(図6)。わが国では、歴史的に組織培養に関する高い知見とノウハウが学術界のみならず民間レベルでも高度に蓄積されているため、本研究成果を活用した技術でそれらをバックアップして産業の活性化につなげたいと考えています。
このように今回の研究成果は、基礎科学にも応用科学にも発展を促すことが予想できます。研究グループはWINDという分子の名前にも、この研究成果が生命科学、特に植物細胞の脱分化研究に新しい風(wind)を吹き込む一因となるようにとの願いを込めています。
発表者
理化学研究所
植物科学研究センター 細胞機能研究ユニット
ユニットリーダー 杉本 慶子(すぎもと けいこ)
基礎科学特別研究員 岩瀬 哲(いわせ あきら)
Tel: 045-503-9570 / Fax: 045-503-9591
独立行政法人産業技術総合研究所
生物プロセス研究部門 主幹研究員
兼 遺伝子転写制御研究グループ 研究グループ長
高木 優(たかぎ まさる)
Tel: 029-861-6717
お問い合わせ先
横浜研究推進部 企画課
Tel: 045-506-9117 / Fax: 045-503-9113
報道担当
独立行政法人理化学研究所 広報室 報道担当
Tel:048-467-9272 / Fax:048-462-4715
独立行政法人産業技術総合研究所 広報部 報道室
Tel: 029-862-6216 / Fax: 029-862-6212
補足説明
- 1.カルス
植物が傷口に作る細胞の塊のこと。癒傷(ゆしょう)組織ともいう。現在では、植物の組織培養で形成される不定形の細胞塊のことを広く示す。植物の体細胞が脱分化して生じると考えられているが、根を再分化しやすい状態や茎葉を再分化しやすい状態をとるなど分化状態は必ずしも均一ではない。これにはカルス細胞内の植物ホルモン(特にオーキシンとサイトカイニン)濃度のバランスや応答性の変化が関与していると考えられる。 - 2.分化、脱分化、再分化
細胞の分化とは、未熟な性質を持った細胞が、より特定の機能を有した細胞に変化することを意味する。例えば、1つの受精卵は細胞分裂をしながらさまざまな機能を持った組織細胞を形成していくが、これは受精卵が分化していく過程である。脱分化は逆に、一度分化し特定の機能を持った細胞がより分化状態の低い細胞に変化することを指す。例えば、形態的にも機能的にも分化し細胞分裂能を失った細胞が、細胞分裂を再開し、再び何かの細胞に分化する能力を獲得した場合、その細胞は脱分化したといえる。脱分化にはさまざまな度合いがあり、少し前の分化状態に戻ることや受精卵のような状態に戻ることのいずれも含む。また、必ずしも細胞の分化の道筋を戻る状態だけに使われるのではなく、がん細胞の形成など、もともと分化の道筋にはない現象も脱分化とみなされる。一度脱分化した細胞が何かに分化するとき、その過程を再分化という。 - 3.シロイヌナズナ
アブラナ科の植物。「ぺんぺん草」で知られるナズナの近縁種である。シロイヌナズナは、植物のモデル生物(普遍的な生命現象の解明に用いられる代表的な生物)の1つ。持っている遺伝子の総量が比較的少なく、発芽から開花し種子が採れるまでの時間が比較的短いことから、被子植物の代表選手として世界中で研究対象になっている。2000年に植物としては初めて全遺伝子の解読が終了したことから、特に分子生物学分野で活躍している。 - 4.転写因子、転写活性化因子、転写抑制因子
特定のDNA配列に結合し遺伝子発現を制御するタンパク質の一群。遺伝子発現を促進する場合と抑制する場合があり、特に前者を転写活性化因子または転写促進因子、後者を転写抑制因子と呼ぶ。 - 5.植物ホルモン、オーキシン、サイトカイニン
植物ホルモンは、植物によって生産され低濃度で植物の生理過程を調節する成長調節物質の総称で、オーキシンやサイトカイニンもこの中に含まれる。サイトカイニンは、オーキシン存在下で細胞分裂や茎葉形成を促進する一群の因子と定義されている。 - 6.組織培養
動物や植物などの組織の一部を取り出して、試験管内の培地で維持したり、培養して増やしたりする技術のこと。植物分野では、種子をつけない植物を大量に増やしたり、ウイルスに感染していない植物を作り出すなど、広く応用されている。 - 7.分化全能性
1つの細胞が個体を構成する全ての細胞種に分化できる潜在能力のこと。植物細胞では、葉肉細胞のプロトプラストや花粉の細胞からカルス化を経て個体を再生できることから、分化全能性を有していると考えられている。 - 8.プロモーターレポーター解析
ある特定の遺伝子のプロモーター領域に、β-グルクロニダーゼ(GUS)遺伝子や緑色蛍光タンパク質(GFP)遺伝子などのレポーター遺伝子をつないで生体内で発現させ、レポーター遺伝子の発現をモニターすることで、その遺伝子がいつ、どの部位で、どんな刺激に応答して遺伝子発現が行われているかなどを観察する解析方法。 - 9.ドミナントネガティブ
ある遺伝子に偶然または人為的に変異が入った時に、その変異遺伝子産物(タンパク質)が正常な遺伝子由来のタンパク質に対して優位(ドミナント)に働いて、かつ正常な遺伝子由来のタンパク質の作用を阻害するとき、ドミナントネガティブ作用が起こるという。 - 10.CRES-T法
CRES-TはChimeric REpressor gene-Silencing Technologyの略。転写活性化因子にSRDXという強い転写抑制能を有する短いアミノ酸配列を付与することで、転写活性化因子を強い転写抑制因子(キメラリプレッサー)に変換する技術。特定の6アミノ酸を付与することで効果があることが明らかになっている。キメラリプレッサーを発現した植物では、ドミナントネガティブな表現型を示す。これは、キメラリプレッサーが内在性の転写因子だけでなく、機能重複する転写因子の転写活性能に優先して標的遺伝子の発現を抑制するためと考えられている。現在植物科学では、強い転写活性化能を付与するVP16と並んで、転写抑制化能を付与するアミノ酸配列としてSRDXが広く用いられている。
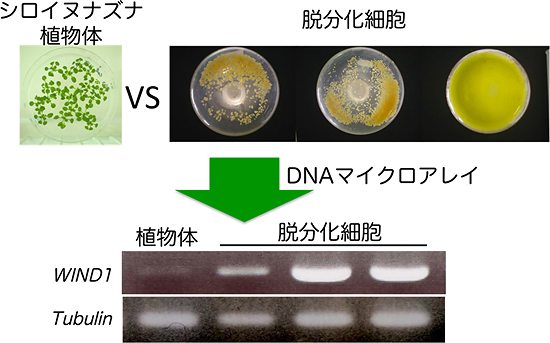
図1 植物の脱分化細胞からWIND1遺伝子を選抜した
モデル植物であるシロイヌナズナを用いて、脱分化した細胞で発現が促進している遺伝子を探索した。植物体と3種類のカルス細胞株に対してDNAマイクロアレイ法による網羅的遺伝子発現解析を行い、分化した植物細胞に比べて3種類のカルス細胞株で共通に発現が促進している遺伝子を選抜した。研究グループはその中から、転写因子の遺伝子に注目し、今回の研究成果に基づいて、この遺伝子をWound INduced Dedifferentiation (WIND)1と名付けた。

図2 WIND1遺伝子は傷口で発現が促進する
WIND1遺伝子のプロモーターレポーター解析(図中の青く染まっている部分が、WIND1遺伝子が発現していると考えられる部位)から、WIND1遺伝子は、傷口に近い細胞(写真左)とそこから生じるカルス(写真右)で特異的に発現が促進することが分かった。

図3 WIND1過剰発現植物はカルスを形成する
WIND1遺伝子をシロイヌナズナ植物体で過剰に発現させると、発現量の大きい植物体ではカルスを形成した。

図4 WIND1発現誘導系カルスはWIND1の誘導を止めると再分化する
WIND1遺伝子の発現誘導系を導入した植物でも、WIND1遺伝子の発現で、植物体や組織片でカルス形成が促進した。このカルスを、誘導剤を含む培地で6カ月継代培養した後に誘導剤を除き、WIND1遺伝子の発現誘導を抑えたところ茎葉や根を再分化した。このことからWIND1で誘導されたカルスは、多分化能を獲得していることが分かった。
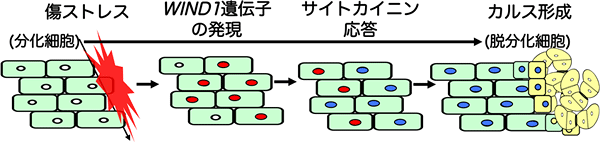
図5 WIND1タンパク質は傷ストレスで誘導される脱分化を制御する
傷口で発現し機能するWIND1タンパク質が、植物ホルモンの一種であるサイトカイニンへの応答を高め、細胞の脱分化を促進することが分かった。
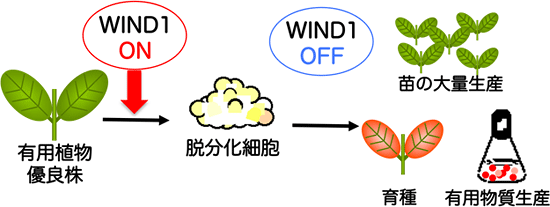
図6 WIND1タンパク質の機能を利用した分子組織培養法の確立を目指す
研究グループは、種苗産業や花卉・園芸産業、植物を用いた医薬品原料生産産業などで、分子スイッチを利用した効率的な脱分化・再分化技術(分子組織培養)の確立を目指している。これにより優良品種の大量栽培、飛躍的な生産性向上に向けた分子育種、有用物質生産のための新形質の付与など、これまでわが国で蓄積された高いノウハウをバックアップすることができると考えている。