2011年3月18日
独立行政法人 理化学研究所
国立大学法人 筑波大学
植物の活性酸素を調節するリン酸化酵素の仕組みを解明
-リン酸化酵素「MAPK」がカルモジュリンと協調して傷害応答を調節-
ポイント
- 定説を覆し、MAPKにはリン酸化反応によらない独自の反応スイッチが存在
- MAPKの1つMPK8は、細胞毒となる過剰な活性酸素蓄積を防ぐ安全弁としても機能
- マラリア原虫などのシグナル伝達経路を特異的に制御する薬剤開発に期待
要旨
独立行政法人理化学研究所(野依良治理事長)と国立大学法人筑波大学(山田信博学長)は、病害虫や干ばつ、塩害など、植物の劣悪環境への応答に関わる活性酸素※1生成のシグナル伝達経路※2に、タンパク質リン酸化酵素の1つ「MAPK(Mitogen-activated protein kinase)※3」が深く関わっていることを世界で初めて解明しました。これは、理研植物科学研究センター(篠崎一雄センター長)機能開発研究グループ/バイオマス工学研究プログラム※4(篠崎一雄プログラムディレクター)バイオマス研究基盤チームの高橋史憲研究員、筑波大学大学院生命環境科学研究科の溝口剛准教授らによる共同研究の成果です。
植物は、自ら移動して育つ場所を選べず、乾燥や水害、日照、病害虫による傷害などの劣悪環境を避けることができないため、独自に環境に適応し、生き抜く能力を備えています。非常に強い酸化力を持つ活性酸素は、植物にとって病原菌を殺す重要な物質となるだけでなく、病害虫による傷害応答、乾燥や塩などへの環境ストレス耐性にも関与しています。一方で、過剰に活性酸素を蓄積すると、逆に細胞毒にもなるため、植物にとってはその厳密な調節が必要不可欠です。しかし、活性酸素をどのように調節して傷害の情報を細胞内で伝達し、必要な応答を引き起こしているのかはよく分かっておらず、長年の課題となっていました。
研究グループは、細胞内のシグナル伝達経路の中で、特にタンパク質のリン酸化反応に着目して研究を行った結果、タンパク質リン酸化酵素の1つ「MAPK」が、植物の傷害応答による活性酸素の生成を調節していることを発見しました。また、これまでの定説を覆し、このTDY型と呼ばれるMAPKの1つMPK8が、リン酸化反応によらない独自のメカニズムで、傷害応答のシグナル伝達反応のスイッチを制御していることを世界で初めて明らかにしました。
今回、活性酸素を調節するシグナル伝達経路を解明したことで、活性酸素の作用を人為的に制御するための基盤を築き上げることができます。また、TDY型MAPKはマラリアなどの原虫にも保存されているため、これら原虫のシグナル伝達を特異的に制御して感染予防に役立つ薬剤を開発するなど、さまざまな応用が期待できます。本研究成果は、米国の科学雑誌『Molecular Cell』(3月18日号)に掲載されるに先立ち、オンライン版(3月17日付け:日本時間3月18日)に掲載されます。
背景
動物や植物、微生物など、酸素を利用してエネルギー代謝を行う生物は、その利用反応の過程で活性酸素を生成しています。この活性酸素は、細胞にとって毒性となる有害物質として知られてきました(図1)。例えばヒトでは、感染、炎症、がん、動脈硬化・糖尿病などの生活習慣病や代謝性疾患など、さまざまな疾病の原因となることが示唆されています。植物でも、乾燥や塩、光などの環境ストレス、病原菌の感染や傷害によって活性酸素を生成し、細胞死を引き起こすことが知られています。植物は移動の自由を持たないため、活性酸素が増えてしまう劣悪環境を避けることはできないうえに、光合成を行って大量の酸素を蓄積しているため、光ストレスを受けると、活性酸素の蓄積が爆発的なものとなってしまいます。従って、植物にとっては、細胞毒となる活性酸素の蓄積をいかに制御するかが、生死を左右する重要な問題です。
しかし近年になると、植物はこの活性酸素をアラーム物質として利用して、傷害の情報を伝達することが明らかになってきました。すなわち活性酸素には、生体分子を損傷させるという悪玉の側面だけでなく、自身の生存に重要なシグナル伝達物質にもなるという善玉の側面もあることが分かってきました。しかし、活性酸素の蓄積制御及びシグナル伝達物質としての利用について、具体的なメカニズムはなかなか解明されず、長い間不明なままでした。
研究手法と成果
MAPKは、動物や植物などの真核生物※5に存在しているタンパク質リン酸化酵素の一種で、タンパク質をリン酸化修飾することで信号を伝達する機能を持ちます。またMAPK自身も、傷害などを含むさまざまなストレスでリン酸化修飾を受け、活性化する特徴があります。
研究グループは、シロイヌナズナのMAPKの中で、植物にしか保存されていないTDY型と呼ぶサブグループに属する8つのMAPK(TDY型MAPK)タンパク質に着目しました(図2a)。これらタンパク質のリン酸化活性を調べた結果、MPK8というタンパク質が、傷害や活性酸素によって活性化することを見つけました。さらに、傷害処理をしたシロイヌナズナの活性酸素蓄積量を測定したところ、MPK8遺伝子を破壊したシロイヌナズナでは、通常の10倍も活性酸素を蓄積することが確認できました。一方、MPK8を過剰に発現させたシロイヌナズナでは、活性酸素を抑制することが確認できたため、MPK8が活性酸素の蓄積を抑制する鍵となるタンパク質であることが分かりました(図2b)。つまりMPK8は、過剰に活性酸素が蓄積しないように安全弁として働き、適切な量の活性酸素をシグナル伝達物質として利用できるように機能していることが判明しました。
そこで研究グループは、MPK8を中心とした活性酸素の蓄積制御及びシグナル伝達のさらなる解明に乗り出しました。はじめに、ゲル内リン酸化反応法※6などの実験を行い、定説通り、上流因子であるMAPKK(MAPK kinase)の1つのMKK3が機能してMPK8はリン酸化され、活性化することが分かりました(図3a)。しかし、MPK8のリン酸化修飾を受けるアミノ酸残基を変換させても、MPK8が活性化することを見いだしました。そこで、Yeast-two hybrid法※7などのさまざまな実験技術を駆使した結果、MPK8の新たな活性化因子となるカルモジュリン※8を発見し、MPK8はリン酸化修飾だけによらず、カルモジュリンによっても活性化することを見いだしました(図3b)。MAPKは、タンパク質リン酸化酵素の1つとして認識されてきましたが、この結果はこれまでのMAPKの定義を覆す新発見となります(図4)。
また研究グループは、TDY型MAPKが植物だけでなく、進化の過程で葉緑体※9の共生※10を経験したマラリアなどの原虫にも存在していることを見いだしました。TDY型MAPKは、ヒトなどの葉緑体を持たない生物には存在しません。つまり、葉緑体を持つ生物は、光合成を行って非常に大量の酸素を体内に持ち、毒となる活性酸素を発生させやすいというリスクを負っているために、進化の過程で、TDY型MAPKを活性酸素の調節機構として利用するようになったと推察されます。
今後の期待
現代社会が抱える食糧問題やエネルギー問題を解決するために、植物バイオマスの育成や増産につながる技術開発が望まれています。今回、植物の劣悪環境への適応に重要な役割を担う活性酸素の調節とそのシグナル伝達の仕組みが分かりました。本研究により、活性酸素を人為的に制御する技術の開発が容易になると考えられます。研究グループは、MPK8を過剰発現した植物は生育に影響無く、一部の除草剤(メチルビオローゲン)などに強くなることを見いだしています(図5)。今回の成果は、活性酸素の量やシグナルを人為的に操作することを可能にし、劣悪環境でもバイオマス増産につながる植物の生育を促進する可能性を秘めています。
また、現在でも熱帯・亜熱帯地域で猛威をふるうマラリア感染は大きな問題となっています。抗マラリア剤などはありますが、副作用があるうえに、マラリアの病原体であるマラリア原虫は成長の過程で劇的に変化するため、有効な予防薬が開発されにくいのが問題の1つとなっています。従って、現状の最大の予防法は、マラリア原虫を媒介するハマダラカ(羽斑蚊)に刺されないよう注意するしかありません。今回研究グループが着目したTDY型MAPKは、マラリアに存在しますが、ヒトには存在しません。マラリアのTDY型MAPKに特異的に作用する薬剤などを今後開発することができると、副作用も少なく有効な抗マラリア剤となる可能性が大いに期待できます。
発表者
理化学研究所
植物科学研究センター 機能開発研究グループ グループディレクター
バイオマス工学研究プログラム バイオマス研究基盤チーム
チームリーダー 篠崎 一雄(しのざき かずお)
研究員 高橋 史憲(たかはし ふみのり)
お問い合わせ先
横浜研究推進部 企画課
Tel: 045-503-9117 / Fax: 045-503-9113
国立大学法人筑波大学
遺伝子実験センター・形質転換植物デザイン研究拠点
センター長補佐・基礎技術研究部門長
生命環境科学研究科 情報生物科学専攻
准教授 溝口 剛(みぞぐち つよし)
Tel: 029-853-6005 / Fax: 029-853-7723
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.活性酸素
生体内のエネルギー代謝の過程で、化学的に活性になった状態の酸素。非常に強い酸化力を持ち、生命を維持するのに必要不可欠な物質である反面、過剰な蓄積は細胞毒となることが知られている。 - 2.シグナル伝達経路
生物がある種の刺激を認識し、その刺激を生体内で連続して伝える過程のこと。シグナル(刺激)の担い手は、シグナル伝達因子という総称で呼ばれる。シグナル伝達因子には、タンパク質やアミノ酸、低分子化合物などさまざまな種類が存在する。タンパク質リン酸化酵素もシグナル伝達因子の一種である。この場合、認識した刺激をリン酸基と呼ばれる化合物に変換し、次のシグナル伝達物質にリン酸基を伝えること(リン酸化)で、生体内での情報をリレーする。 - 3.MAPK(Mitogen-activated protein kinase)、MAPKK(MAPK kinase)
タンパク質リン酸化酵素の一種で、環境ストレス応答や、病原菌応答、細胞増殖などのシグナル伝達に関わる。MAPKKはMAPKにリン酸基を付加し、さらに次の因子にリン酸基を付加するという一連のリン酸化リレーが、リン酸化酵素の定義となっている。シロイヌナズナには20個の MAPK遺伝子が存在し、そのうちの8個の遺伝子がTDY型と呼ばれるタイプに属する。 - 4.バイオマス工学研究プログラム
バイオマス工学研究プログラム(Biomass Engineering Program:BMEP)は、2010年4月、理研に発足した新しい研究プログラム。二酸化炭素の資源化に向け、植物の光合成によりセルロースなどのバイオマスを増産するための研究を推進し、生産した植物バイオマスを原料としてバイオプラスチックなどの新バイオ素材を創る新たな技術を確立することにより、“グリーン・イノベーション”の創出、つまりは社会知の形成に貢献することを最大の目的としている。 - 5.真核生物
生体を構成する細胞の中に、ゲノムなどが入った細胞核と呼ばれる構造を持つ生物を指す。動物や植物、菌類、原生生物などが属する。 - 6.ゲル内リン酸化反応法
SDS-PAGEゲルに基質ポリペプチドを固定化して埋めこみ、そこにタンパク質リン酸化酵素を電気泳動して、ゲルごとリン酸化反応を行わせる手法。リン酸化酵素の活性量とともに、その分子量が同時に分かる。 - 7.Yeast-two hybrid法
出芽酵母 Saccharomyces cerevisiaeを用いて、2つのタンパク質の相互作用を調べる実験。結合能を調べようとする2つのタンパク質を出芽酵母の中に導入し、酵母が生育することで結合の有無を確認することができる。 - 8.カルモジュリン
さまざまな生物に存在するカルシウム結合タンパク質で、いろいろな細胞内シグナル伝達経路に関わっていることが知られている。カルモジュリンはカルシウムと結合したのち、目的のタンパク質と結合することで、カルシウムを使ったシグナル伝達因子として働く。 - 9.葉緑体
細胞内に存在する細胞小器官の1つである。光合成を行う器官としてよく知られているが、植物細胞では、その他に窒素代謝やアミノ酸合成など代謝の重要な中心となっている。 - 10.共生
二種の生物間に成り立つ捕食関係、寄生関係、競争関係、共生関係を意味する。ここでは細胞内共生の意を指し、光合成を行う真正細菌などを細胞内に共生させたことが、植物などの真核光合成生物の起源であると考えられている。
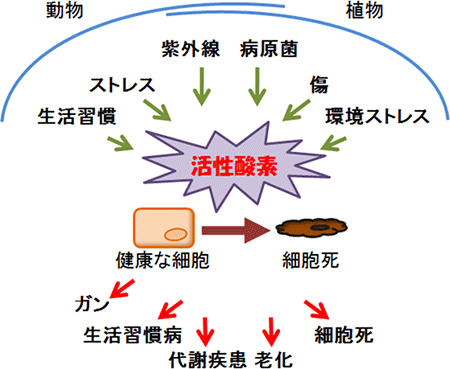
図1 活性酸素はさまざまなストレスによって生成する
ストレスや劣悪環境などにさらされると、動物や植物の生体内では、活性酸素が増加する。過剰な量の活性酸素は、健康な細胞を死滅させる毒となり、さまざまな疾病や老化、細胞死の原因になることが報告されている。
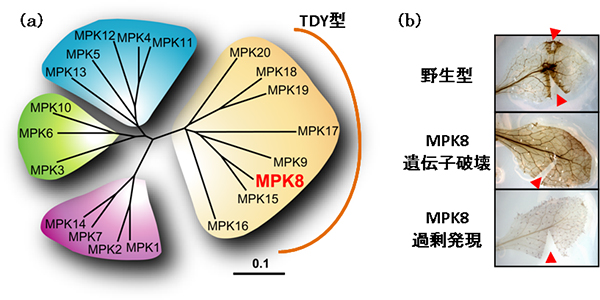
図2 TDY型MAPKの1つMPK8は活性酸素の蓄積を制御する
- (a) シロイヌナズナのMAPKは20遺伝子存在し、大きく4つのサブグループに分けられる。そのうちMPK8が属するTDY型MAPKは8遺伝子が含まれ、最も大きなサブグループを形成する。TDY型MAPKは、植物など葉緑体との共生を経験した生物にだけ存在する。
- (b) 傷害処理(赤▲は切れ込みを示す)2時間後の活性酸素の蓄積を観察した実験。茶褐色に変色した組織に活性酸素が蓄積していることを示す。野生型では、切れ込みを入れた周辺に活性酸素が蓄積しているのに対し、MPK8遺伝子破壊植物では、すでに葉全体に蓄積しているのが分かる。一方、MPK8過剰発現植物では活性酸素の抑制が認められる。これによりMPK8が活性酸素の蓄積を制御していることが判明した。
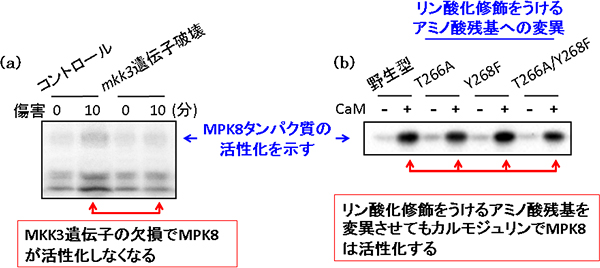
図3 MKK3とカルモジュリンはMPK8を活性化する
- (a)
MKK3によるMPK8の活性化をゲル内リン酸化法で調べた実験
傷害処理10分後のMPK8のリン酸化活性を調べた結果(青矢印)、コントロールで観察たMPK8の活性は、MKK3遺伝子を欠損させると観察できなくなった。これにより、MKK3はMPK8に情報を伝達していることが分った。 - (b)
カルモジュリンによるMPK8の活性化を試験管内リン酸化法で調べた実験
通常、タンパク質リン酸化酵素は、リン酸化修飾をうけるアミノ酸残基を変換させると、リン酸基を受け取れることができなくなるため、リン酸化活性がなくなる。しかしカルモジュリンは、MPK8のアミノ酸変換に関わらず、MPK8を活性化することが分かった。これにより、MAPKの定義を覆す新しい活性化のメカニズムを発見することにつながった。
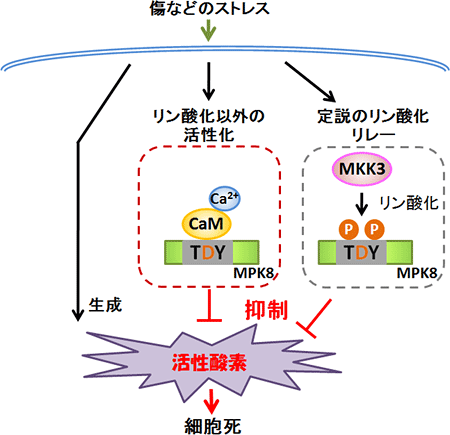
図4 研究グループが明らかにしたMAPKによる活性酸素の制御メカニズム
傷などのストレス刺激を受けると、活性酸素を生成するシグナルが動く。一方、MKK3はリン酸化リレーを使ってMPK8を活性化し、活性酸素の生成を抑制する。さらにカルモジュリンはMPK8をリン酸化しないで活性化し、活性酸素の生成を抑制する。MPK8は過剰に活性酸素が蓄積しないように安全弁として働き、適切な量の活性酸素をシグナル伝達物質として利用するように働いていることを明らかにした。MAPKはタンパク質リン酸化酵素の1つとして認識されてきたが、これまでのMAPKの定義を覆し、リン酸化を受けない新たな活性化メカニズムがあることを発見した。

図5 MPK8は薬剤などによる酸化ストレスに耐性を示す
メチルビオローゲン処理後の植物を観察した実験で、MPK8遺伝子破壊植物は、はやくに細胞死が起こるため、組織が黄化しすぐに死滅してしまう。一方MPK8過剰発現植物は、耐性を示し細胞が死なない。