ポイント
- タンパク質脱リン酸化酵素のメチル化が、ブラシノステロイド受容体の量を制御
- ホルモンと結合した活性型受容体は分解、結合していない受容体は再利用
- ロイシンリッチリピートを持つ受容体の負の調節機構を初めて明らかに
要旨
独立行政法人理化学研究所(野依良治理事長)は、植物ホルモン※1の1つであるブラシノステロイド※2の受容体BRI1の量は、ブラシノステロイドの量に応じて調節されるという、情報伝達※3の新たな機構を発見しました。これは、理研植物科学研究センター(篠崎一雄センター長)生長制御研究グループの神谷勇治グループディレクター、グアン・ウー(Guang Wu, JSPS fellow)とカリフォルニアSalk研究所ジョアン・コーリー(Joanne Chory)教授との共同研究の成果です。
ブラシノステロイドは、植物の茎の生長や葉の大きさの制御など、植物の生長と分化を調節する生長ホルモンとして知られています。ブラシノステロイドが合成できない、または受容できない突然変異体は、野生型に比べて茎や葉が極端に矮(わい)化します。ブラシノステロイドの受容体BRI1は、細胞膜に存在するロイシンリッチリピート※4を持つ膜貫通型のセリン/トレオニンキナーゼ型受容体で、細胞膜と細胞内との間を循環することが知られています。しかし、細胞内で受容体BRI1の量がどのように調節されているかは不明のままでした。研究グループは、矮性を示すbri1突然変異体から野生型のように草丈が戻る突然変異体を探索して、その原因遺伝子SBI1を同定しました。さらに、この遺伝子がコードするSBI1タンパク質が、タンパク質脱リン酸化酵素2A(PP2A)※5を特異的にメチル化するロシインカルボキシルメチル転移酵素※6であることを発見しました。メチル化されたPP2Aは立体構造が変化して、ブラシノステロイドと結合してリン酸化した活性型BRI1との親和性が高まり、BRI1タンパク質を脱リン酸化すると同時に、タンパク質分解酵素による分解へと導くことが明らかになりました。一方、ブラシノステロイドと結合していない細胞内にある不活性型BRI1は、タンパク質分解酵素によって分解されずに、再び細胞膜に運ばれて受容体として再利用されることが判明しました。これは、これまで知られていなかったロイシンリッチリピートを持つ受容体タンパク質の活性依存的な負の調節機構です。
膜貫通型のセリン/トレオニンキナーゼ型受容体は広く生物に存在し、ホルモンなどの活性分子の情報伝達に関わっています。この受容体の異常は、ヒトの病気とも関連しているため、今回の成果は、植物、動物の生命現象に新たな知見をもたらすと期待されます。
本研究成果は、米国の科学雑誌Scienceの姉妹誌『Science Signaling』(オンライン版(5月10日付け:日本時間5月11日)に掲載されます。
背景
植物ホルモンのブラシノステロイドは、植物の茎の生長や葉の大きさの制御、ストレス抵抗性の向上など、植物の生長と分化のさまざまな段階で生理作用を発揮します。ブラシノステロイドが生合成できない、または受容できない突然変異体は、ブラシノステロイドの情報伝達がうまく行かず、野生種に比べて茎や葉が極端に矮化します(図1、bri1-5突然変異体)。理研基幹研究所とSalk研究所の研究グループはこれまでに、ブラシノステロイドの受容体BRI1がロイシンリッチリピートを持つ膜貫通型のセリン/トレオニンキナーゼ型受容体であることを明らかにしてきました (Cell 1997、Science 2000、Nature 2001)。ブラシノステロイドの情報伝達に関わる遺伝子の中に、BRI1受容体の下流で働く遺伝子としてBIN2があります。BIN2タンパク質はショウジョウバエのSHAGGYや哺乳類のGSK3と相同な細胞質に存在するキナーゼです。BRI1のような膜貫通型のセリン/トレオニンキナーゼ型受容体からBIN2のような細胞質型キナーゼに、どのような方法でシグナルが伝わるのか、またBRI1タンパク質が植物細胞内でどのように量的に調節されているかは分かっていませんでした。ロイシンリッチリピートを持つ膜貫通型のセリン/トレオニンキナーゼ型受容体は生物に共通して存在し、この欠損はヒトの病気とも関連しています。このタイプの受容体はシロイヌナズナには600以上もあり、これらが結合する基質もほとんど分かっていません。従って、これらの受容体の調節機構を明らかにすることは、生命現象の基礎を理解する上で非常に重要となります。
研究手法と成果
(1)BRI1の変異を回復させるSBI1遺伝子の同定
シロイヌナズナのbri1突然変異体には、遺伝子上の変異の場所により極めて矮性の強い突然変異体や矮性の弱い突然変異体など、数多くの種類があります。完全欠損突然変異体は不稔になるために種子を得られませんが、BRI1タンパク質の量が野生種に比べて10分の1ほどに少なくなったbri1-5突然変異体は、矮性を示しながらも種子を作ることができます。このbri1-5種子に突然変異を起こす化学試薬を処理して、約20,000の種子から矮性が回復する復帰突然変異体を探索しました。その結果、光条件下で育てると野生種と同等までに草丈が回復する変異体が得られ(図1、sbi1/bri1-5二重突然変異体)、原因遺伝子としてSBI1遺伝子を同定しました。さらにSBI1遺伝子単独の突然変異体を作製したところ、驚いたことに野生種より草丈が大きくなりました(図1、sbi1突然変異体)。この結果は、SBI1遺伝子がブラシノステロイドの情報伝達の負の制御因子である可能性を示しています。
(2)ブラシノステロイド受容体BRI1の調節モデルの検討
細胞膜から細胞内に運ばれたBRI1が、再び細胞膜に送られて再利用されるか、それとも細胞内でタンパク質分解酵素によって分解されるかは、ブラシノステロイドの情報伝達の調節にとって重要といえます。これまでの研究から、BRI1は、常に細胞膜と細胞内との間を循環していることが知られています。sbi1遺伝子のポジショナルクローニング※7を行ったところ、SBI1遺伝子は、シロイヌナズナのロイシンカルボキシルメチル転移酵素SBI1をコードしていることが明らかになりました。この酵素の標的はタンパク質脱リン酸化酵素2A(PP2A)で、このPP2Aはブラシノステロイドが結合してリン酸化された活性型BRI1のセリン/スレオニン残基の側鎖のリン酸基を脱リン酸化し、分解へと導く働きを持っています。
PP2Aが欠損した突然変異体rcn1では、野生型に比べてBRI1とブラシノステロイドの量が増加します。sbi1突然変異体ではブラシノステロイドによってリン酸化された活性型BRI1の量は増加しますが、BRI1と相互作用をするもう1つの膜貫通型のキナーゼBAK1は全く変化しませんでした。これは、PP2AがBRI1の分解に特異的に関与することを意味しています。また、SBI1のmRNAの発現量はブラシノステロイドの量に応じて増加しました。すなわち、細胞膜でBRI1がブラシノステロイドを受容すると、ブラシノステロイドの量に応じてSBI1が合成されることが分かりました。
これらの結果から、SBI1が、PP2Aをメチル化すると、PP2Aの立体構造が変化して、リン酸化された活性型BRI1との親和性が高まり、その結果、活性型BRI1の脱リン酸化と分解が起こる、という調節機構が明らかとなりました(図2)。
(3)ホルモンの量に応じた負の調節機構の発見
bri1突然変異体の中には、BRI1のブラシノステロイドの結合部位に変異が入ったもの、キナーゼ活性を持つ部位に変異が入ったもの、BRI1タンパク質全体を作ることができないものなど、突然変異体の性質が明らかになっているものがあります。これらの突然変異体を使ってBRI1の細胞内での安定性を調べました。その結果、ブラシノステロイドが結合した活性型BRI1はタンパク質分解酵素で選択的に分解され、細胞内にあるリン酸化されていない不活性型BRI1は分解されずに細胞膜に運ばれて受容体として再利用されることが分かりました。これは、ブラシノステロイドが結合した活性型BRI1が選択的に分解されることにより、ブラシノステロイドの総シグナル量が低下して、情報伝達が終了することを示しています。これにより、ブラシノステロイドの量に応じた受容体の分解による負の調節機構存在が初めて明らかになりました。
今後の期待
この研究により、BRI1の量がホルモンとの結合依存的に調節され、活性化されていない未使用の受容体は誤って分解されることなく、再利用されることが分かりました。この調節に関わる酵素が、ロイシンカルボキシルメチル転移酵素SBI1であることも判明しました。植物ホルモンの情報伝達には多くの転写因子が関与していますが、これまで実用化され農業で利用されている植物生長調節剤は、ホルモン自身かその誘導体、またはホルモン生合成酵素の阻害剤などです。ホルモンの情報伝達に関わる転写因子は複数の標的を持つために、実用化された調節剤はほとんどありません。今回のブラシノステロイドの情報伝達に関わる突然変異体の探索では、転写因子ではなく、メチル基転移酵素のSBI1が単離されてきました。従って、この酵素の選択的な阻害剤は新しい植物生長調節剤の可能性があります。
ロイシンリッチリピートを持つ膜貫通型のセリン/トレオニンキナーゼ型受容体は、広く生物に存在し、いろいろな物質の受容体として働いています。ヒトではTGF-β※8が良く知られていて、TGF-βの欠損は病気と深く関わっています。また、TGF-βを脱リン酸化する酵素PP2Aの機能異常は、がんやアルツハイマー病との因果関係があることが知られており、植物におけるPP2Aを介したブラシノステロイドの情報伝達の研究は、動物を含めた生命現象を理解する上で重要といえます
植物には、多くのロイシンリッチリピートを持つ膜貫通型のセリン/トレオニンキナーゼ型受容体が存在します。それらの情報伝達を調べることにより、今回明らかになった調節機構が生物に共通するものかどうかを突き止めることができます。それにより、植物と動物の分子進化についても新たな知見を得られると期待されます。
発表者
理化学研究所
植物科学研究センター 生長制御研究グループ
グループディレクター 神谷 勇治(かみや ゆうじ)
Tel: 045-503-9661 / Fax: 045-503-9662
お問い合わせ先
横浜研究推進部
Tel: 045-503-9117 / Fax: 045-503-9113
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.植物ホルモン
植物自身が作り出し、微量で作用する生理活性物質・情報伝達物質。植物に普遍的に存在し、その物質の化学的本体と生理作用が明らかにされたもの。オーキシン、エチレン、ジベレリン、アブシシン酸、サイトカイニン、ジャスモン酸、ブラシノステロイド、ストリゴラクトン、サリチル酸、ペプチドホルモンなどがある。 - 2.ブラシノステロイド
花粉から最初に発見された植物ステロイドホルモン。細胞伸長、細胞分裂、屈曲、栄養分の運搬、木部分化、老化促進、抗ストレスなどの作用を持つ。カンペステロールから生合成される。 - 3.情報伝達
ホルモンなどのシグナル分子が、細胞で受容されて、細胞内を移動し、最終的には核で遺伝子の発現を調節して効果を発揮するまでの過程を指す。シグナル分子としては細胞外のシグナル媒体と細胞内のシグナル媒体がある。 - 4.ロイシンリッチリピート
タンパク質の構造モチーフの1つ。20~30残基のアミノ酸の繰り返し配列で、疎水性のロイシンの割合が多い。ブラシノステロイドの受容体BRI1は、この構造を細胞膜の外側に持つキナーゼ型の受容体である。 - 5.タンパク質脱リン酸化酵素2A(PP2A)
セリン/トレオニン残基の側鎖のリン酸基を加水分解するタイプ2型の酵素。細胞周期や細胞のシグナル伝達に関わっている。A, B, Cの3つのサブユニットからできている。 - 6.ロイシンカルボキシルメチル転移酵素
タンパク質脱リン酸化酵素2Aのロイシンのカルボキシル基をメチル化する酵素。 - 7.ポジショナルクローニング
ファンクショナルクローニングに対して、染色体上の異常部位(ポジション)から遺伝子の解析を行うこと。ゲノム情報の知られているモデル生物の場合は、異常部位を見つけるだけで遺伝子を特定化することができる。 - 8.TGF-β
トランスフォーミング成長因子。増殖因子の1つで、組織発生、細胞分化、胚発生において重要な役割を持つ。腎臓、骨髄、血小板など、ほとんど全ての細胞で生産されている。

図1 光のもとで育てたブラシノステロイドの突然変異体
bri1-5:bri1-5ブラシノステロイド受容体突然変異体
sbi1/bri1-5:復帰突然変異体(sbi1/bri1-5二重突然変異体)
WT:野生種
sbi1:sbi1突然変異体
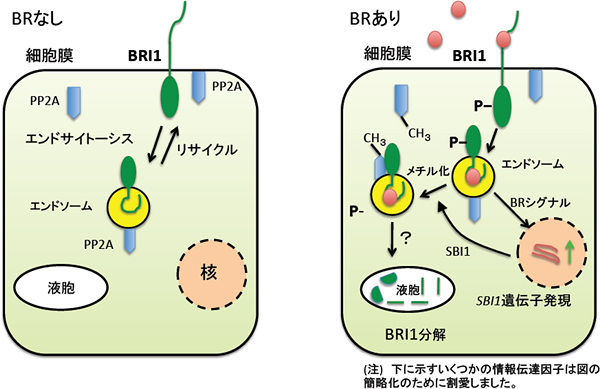
図2 ブラシノステロイド受容体BRI1と総ブラシノステロイド量の調節モデル
- (左)ブラシノステロイド(BR)が無い場合、BRI1は細胞膜と小胞(エンドソーム)の間を循環している。
- (右)ブラシノステロイド(BR)を受容した場合、BRI1は細胞膜上でBRと結合することによりリン酸化され、活性型となる。この活性型BRI1はBRのシグナルを核に伝え、SBI1を含めた遺伝子の発現を誘導する。SBI1は標的酵素のPP2Aをメチル化して、それにより活性型BRI1の脱リン酸化が起こる。脱リン酸化されたBRI1はおそらく液胞内で分解され、BRシグナルが消失する。図中の「?」は、いまだ証明されていない過程を示している。このモデルでは、BRシグナルが細胞膜上だけで受容されるように示しているが、細胞内のエンドソーム膜上でもBRシグナルの受容が起こる可能性は否定できない。なお、ここでは、BRの情報伝達に関わるBAK1, BSK1, BKI1, BIN2, BES1, BZR1, BSU1は省略している。