2012年5月29日
独立行政法人 理化学研究所
栄養素を運ぶタンパク質「NRT1.2」が植物ホルモンも運ぶことを発見
-植物ホルモン受容体の複合体形成を利用して、その輸送体を網羅的に探索-
ポイント
- NRT1.2を欠損すると花茎の気孔が約1.4倍大きく開き、表面温度が低下する
- 発見されていない他の植物ホルモンの輸送体探索にも適用可能
- NRT1.2の機能を改変し、乾燥ストレス耐性・休眠性と生長の同時制御に期待
要旨
理化学研究所(野依良治理事長)は、根から吸収した栄養素を運ぶタンパク質「NRT1.2」が、植物ホルモン※1のアブシシン酸※2の輸送体※3であることを、シロイヌナズナを用いた実験で発見しました。これは、理研植物科学研究センター(篠崎一雄センター長)適応制御研究ユニットの瀬尾光範ユニットリーダー、菅野裕理テクニカルスタッフ、生長制御研究グループの神谷勇治グループディレクター、植物ゲノム機能研究グループの松井南グループディレクターと、首都大学東京大学院理工学研究科生命科学専攻植物ホルモン機構研究室の小柴共一教授らによる共同研究グループの成果です。
植物ホルモンの働きは植物体内に存在するホルモンの量に依存し、その量は合成される量と分解される量のバランスによって決まります。また、植物ホルモンは作られた場所から移動し、局所的に分布が変化します。多くの植物ホルモンの合成と分解に必要な酵素や遺伝子は見つかっていますが、移動に関わる因子、つまり輸送体はほとんど見つかっていません。
共同研究グループは、気孔の閉鎖に必要な植物ホルモンであるアブシシン酸(ABA)の受容体が、ABAを認識すると特定のタンパク質と複合体を形成することをセンサーとして利用し、ABA輸送体を網羅的に見つけ出す方法を開発しました。これを使い、栄養素の硝酸の輸送体であるタンパク質「NRT1.2」が、ABAの輸送体として働くことを発見しました。植物体内でこれら2つの物質輸送を同時に変化させることで、ストレス耐性付与と硝酸輸送を変化させ、より高度な生長制御が可能になると考えられます。
今回開発した受容体をセンサーとする輸送体の発見法は、植物の生長を制御するジベレリンや傷害・病害防止に必要なジャスモン酸など、まだ輸送体が発見されていない植物ホルモンにも応用が可能です。これらの植物ホルモン輸送体の機能を改変することで、農作物などの生産性の向上が可能になると期待できます。
本研究成果は、米国科学アカデミー紀要『Proceedings of the National Academy of Science of the United States of America』オンライン版に5月28日の週に掲載されます。
背景
ホルモンとは、動物体内のある特定の場所(器官)で作られた後、体液(血液)を介して運ばれて、離れた場所(器官)で働く生理活性物質の総称です。植物にも同様な生理活性物質が存在し、これらは動物のホルモンとは区別されて、「植物ホルモン」と呼ばれます。一般に植物ホルモンは、①植物体内で作られ②10-6M(モーラー:モル濃度)以下の低濃度で作用し③植物種を超えて普遍的に存在する低分子化合物と定義されます。進化論で有名なダーウィンが発見したオーキシンは、植物体内を決まった方向に輸送されることが知られています。しかし、他の植物ホルモンについては、どこで作られ、どのように移動し、どこで作用するかがほとんど分かっていません。
アブシシン酸(ABA)は、葉の裏などにある気孔の閉鎖、ストレス耐性遺伝子の発現誘導、種子休眠の誘導などに必要な植物ホルモンです。2004年頃には、ABAは水分や有機物の通路となる維管束組織で合成された後、気孔を形成する孔辺細胞へ輸送されると考えられていました。しかし、それを明らかにするにはABAを維管束組織から孔辺細胞に運ぶ輸送体の同定が不可欠でした。
研究手法と成果
植物ホルモンは、特定の受容体に認識されて初めてその作用を発揮します。ABA受容体はABAを認識するとタンパク質脱リン酸化酵素PP2C(protein phosphatase 2C)※4と複合体を形成します。その結果、PP2Cによるタンパク質リン酸化酵素SnRK2(SNF1-related protein kinase 2)※5に対する抑制が無くなり、SnRK2は活性化します。これによって転写因子などを含む標的タンパク質がリン酸化され、下流の情報伝達系が活性化されます(図1)。この複合体形成は、酵母two-hybrid系※6を用いることで簡便に検出可能です。
そこで共同研究グループは、この複合体形成を酵母細胞内のABA濃度を感知する「センサー」とし、酵母two-hybrid系を応用することで、輸送体を網羅的に見つけ出す方法を開発しました。培地中のABA濃度が低い場合には、酵母細胞内のABA濃度も低いため、ABA受容体とPP2Cの複合体形成が起きません(図2a)。しかし、ABA輸送体を持つ酵母はABAを細胞内に取り込むことができるため、ABA濃度が低い培地中でも十分に酵母細胞内のABA濃度が上昇し、受容体とPP2Cの複合体形成が起きます(図2b)。実験の結果、硝酸の輸送体として知られている「NRT1.2」タンパク質が、ABA輸送体として働くことが分かりました。
NRT1.2を欠損させた変異体を調べたところ、その花茎では気孔の開き幅(気孔開度)が野生型と比べて約1.4倍大きくなり、気化熱による表面温度が低下したことを確認しました(図3)。つまり、ABAが孔辺細胞に正常に輸送されず、気孔が閉じにくくなったことが示されました。このことから、NRT1.2が気孔を閉鎖して乾燥耐性を獲得するために重要な役割を果たしていると分かりました。
今後の期待
ABA輸送体の機能を改変することで、乾燥などのストレス耐性の付与や休眠性の制御など、農業への応用が可能になると期待できます。同時に、硝酸の輸送を変化させることができると、より高度な生長制御が期待できます。
また、今回開発したABA輸送体の発見方法は、植物の生長を制御するジベレリンや傷害・病害防止に有効なジャスモン酸など、他の植物ホルモンにも適用可能です。今後、新たな植物ホルモン輸送体を見つけていくことで、さまざまな環境での植物生長の最適化が可能になると期待できます。
原論文情報
- Yuri Kanno, Atsushi Hanada, Yasutaka Chiba, Takanari Ichikawa, Miki Nakazawa, Tomokazu Koshiba, Yuji Kamiya and Mitsunori Seo “Identification of an abscisic acid transporter by functional screening using the receptor complex as a sensor”. Proceedings of the National Academy of Sciences of the United States of America, 2012, doi/10.1073/pnas.1203567109
発表者
理化学研究所
植物科学研究センター 適応制御研究ユニット
チームリーダー 瀬尾 光範(せお みつのり)
お問い合わせ先
横浜研究推進部 企画課
Tel: 045-503-9117 / Fax: 045-503-9113
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.植物ホルモン
現在までにオーキシン、ジベレリン、サイトカイニン、エチレン、アブシシン酸、ブラシノステロイド、ジャスモン酸、サリチル酸、ストリゴラクトンの9種が知られている。ペプチド性の生理活性物質は「ペプチドホルモン」として区別される。 - 2.アブシシン酸(ABA)
カロテノイドを前駆体として合成されるテルペン類化合物の一種。ABAを合成できない変異体では、気孔の閉鎖ができず、過剰な蒸散のためしおれやすい。ABAは、乾燥などのストレスに抵抗するため活発に合成され、ストレス耐性遺伝子の発現を誘導する。ABAは、種子休眠の誘導や発芽の阻害にも関与する。コムギなどの穀類種子が収穫前に母体植物上で発芽してしまう穂発芽は、ABAの内生量の低下やABAに対する感受性の低下と関連していることが多い。 - 3.輸送体
生体膜に存在するタンパク質で、水素イオンなどの一次エネルギー勾配を利用して物質輸送を行う。トランスポーターとも呼ばれる。 - 4.PP2C(protein phosphatase 2C)
タンパク質脱リン酸化酵素の一種で、植物のABA応答を負に制御する。1990年代に、ABAへの感受性が弱まったシロイヌナズナ変異体の原因遺伝子ABI1として最初に同定された。シロイヌナズナには9つの類似した遺伝子が存在する。 - 5.SnRK2(SNF1-related protein kinase 2)
タンパク質リン酸化酵素の1種で、シロイヌナズナには9つのファミリーが存在する。そのうち、SRK2D、E、Iの3つがABAによって強く活性化し、ABA応答を正に制御する。 - 6.酵母two-hybrid系
酵母細胞内で2種類のタンパク質の相互作用を検出する方法。通常、一方のタンパク質に特定のDNA結合配列を付加し、もう一方のタンパク質に転写活性化領域を付加する。2種のタンパク質が複合体を形成した場合には、複合体が1つの転写活性化因子として機能し、下流の遺伝子発現を活性化する。選択培地上で酵母が生育可能か否かで、複合体形成の有無を判断できる (図2参照)。
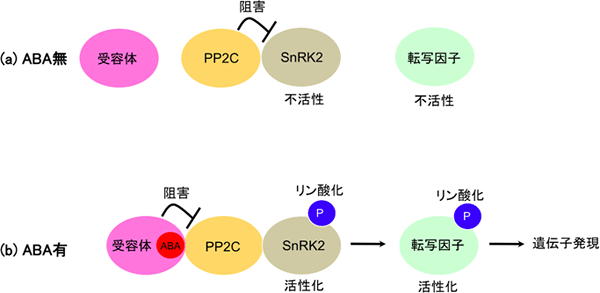
図1 アブシシン酸の受容と情報伝達の初期反応
- (a) アブシシン酸(ABA)がない時(ABA濃度が低い時)は、タンパク質脱リン酸化酵素PP2Cがタンパク質リン酸化酵素SnRK2の活性を抑制する。それにより、下流の転写因子などが不活性化状態になる。
- (b) ABAがある時(ABA濃度が高い時)は、受容体がABAの認識に伴いPP2Cと複合体を形成しその活性を阻害する。これによりSnRK2が活性化し、下流の転写因子などを活性化させることで、ABAに対する生理応答が引き起こされる。
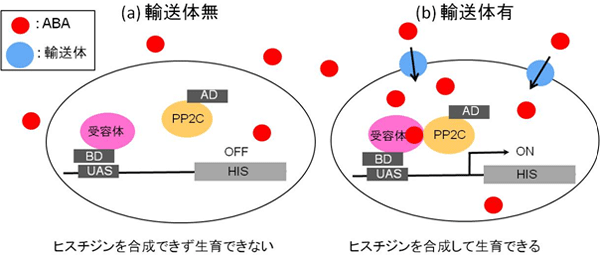
図2 酵母two-hybrid系を利用した輸送体の同定
細胞の生育に欠かせないアミノ酸の1つであるヒスチジンを合成できない酵母細胞を用いた実験。酵母細胞内でヒスチジン合成遺伝子(HIS)の上流域(UAS)に結合するDNA結合領域(BD)を付加したABA受容体と、転写活性化領域(AD)を付加したPP2Cを発現させる。培地中のABA濃度を低くして、ヒスチジンの合成を起こりにくくする。
- (a) 輸送体がない場合、ABA受容体とPP2Cの複合体形成は起きない。その結果、酵母細胞はヒスチジンを合成できず生育できない。
- (b) 輸送体がある場合、細胞膜外から内へABAを取り込むため酵母細胞内のABA濃度が上昇する。その結果、ABA受容体とPP2Cが複合体を形成し、付加されたADのはたらきにより、HIS遺伝子が発現、ヒスチジン合成をするため、酵母は生育可能になる。
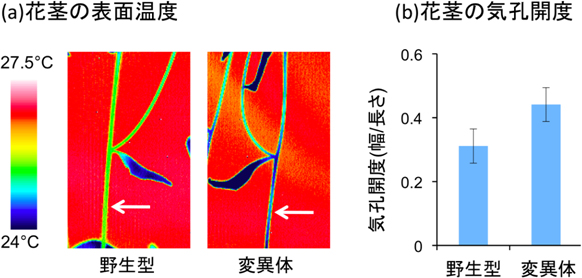
図3 野生型とNRT1.2を失った変異体の温度変化と気孔開度
- (a) 野生型と変異体の花茎の表面温度を赤外線カメラで観察。矢印は花茎を指す。変異体は野生型より表面温度が低いことが分かる。
- (b) 野生型と変異体の花茎の表面に存在する気孔の開度(幅/長さ)を測定。変異体の気孔が、野生型に比べて約1.4倍開いていることが分かる。