2012年10月25日
独立行政法人理化学研究所
タンパク質合成を促進するアンチセンスRNAを初めて発見
-翻訳促進に関わるRNA上の特殊な配列とそのメカニズムを解明-
ポイント
- Uchl1遺伝子のアンチセンスRNAが、タンパク質の合成を促す
- タンパク質発現の増大にアンチセンスRNA上の特殊な配列「SINEB2」が関与
- タンパク質をコードしないncRNAで新たな機能を発見
要旨
理化学研究所(野依良治理事長)は、これまで生体内におけるタンパク質合成を阻害すると考えられていたアンチセンスRNA※1の中に、タンパク質合成を促進する機能も持つものがあることを発見しました。これは、理研オミックス基盤研究領域(林崎良英領域長)LSA要素技術開発グループゲノム機能研究チームのピエロ カルニンチチームリーダーらと、イタリアのInternational School of Advanced Studies(SISSA)との共同研究グループによる成果です。
一般にアンチセンスRNAは、メッセンジャーRNA(mRNA)※2からタンパク質を合成(翻訳)する機能を阻害すると知られていますが、最近では、ゲノムの転写やmRNA(センスRNA)の安定性を制御する機能についても報告されています。
共同研究グループは、アンチセンスRNAのさらなる機能を明らかにするため、神経変性疾患などに関与し「センスRNA-アンチセンスRNA」のペアが発現する遺伝子Uchl1に着目し、その翻訳機構を調べました。その結果、リボゾーム※3との親和性が高く、かつ特殊な配列を持つアンチセンスRNA「SINEB2」が、これと相補的配列を持つセンスRNAとの相互作用を促進し、タンパク質合成を促進させることが分りました。このアンチセンスRNAは、国際研究コンソーシアム「FANTOM※4」で見いだされたノンコーディングRNA(ncRNA)※5の1つです。細胞内には、mRNAなど機能あるRNAの他に、アンチセンスRNAなどタンパク質をコードしないRNA(ncRNA)が大量に存在します。これまでの定説では、ncRNAのほとんどはジャンクと考えられていました。しかし、今回の成果は、ncRNAにも生命現象維持に役立つ機能がある可能性を示しています。本研究成果は、英国の科学雑誌『Nature』オンライン版(10月14日付け:日本時間10月15日)に掲載されました。
なお、カルニンチチームリーダーらは、今回の成果をもとに社会貢献するためにベンチャー会社「トランスサイン テクノロジーズ株式会社(TransSINE Technologies Co.Ltd)」を設立しました。さまざまなニーズに応じた目的のタンパク質合成を促進するアンチセンスRNAを開発し、基礎研究から医療まで幅広い分野での貢献を目指します。理研は、トランスサイン テクノロジーズ株式会社を理研ベンチャー※6に認定しており、本技術の迅速な実用化と普及を支援します。
背景
哺乳類のゲノムのほとんどの領域はRNAに転写されて、タンパク質をコードするmRNAや、タンパク質をコードしないncRNA、短鎖酸在反復配列(SINE)※7のようにジャンクと呼ばれる同じ配列が反復しているだけのものなどが生成されています。ncRNAの多くは核内にとどまることが知られていますが、その全ての機能は解明されていません。ncRNAのうち塩基配列が100~200bp以上のアンチセンスlncRNAと呼ばれるものは、これと相補的な配列を持つmRNA(センスRNA)と結合して、その翻訳を阻害することがよく知られています。一方、近年の研究では、ゲノムの転写や、mRNAの安定性を制御する可能性などが報告されています。
研究手法と成果
共同研究グループは、アンチセンスRNAのさらなる機能を明らかにするため、脳機能や神経変性疾患に関与して脳内で「センスRNA-アンチセンスRNA」のペアが発現している遺伝子Uchl1に着目し、その転写翻訳機構を調べました。マウスの細胞からUchl1のアンチセンスlncRNA(アンチセンスUchl1)の配列を解析したところ、Uchl1のセンスRNA(センスUchl1)と相補しない領域に2つのSINE(SINEB2、Alu)が見つかりました。また、アンチセンスUchl1の5’末端に、センスUchl1の5’末端と結合する配列を有しており(図1)、このアンチセンスUchl1のゲノムにおける配列が哺乳類の遺伝子に共通に存在するものということも分かりました。
センスUchl1の発現が見られるマウスの細胞で、アンチセンスUchl1を発現させたところ、センスUchl1量に変化はなかったものの、UCHl1タンパク質の合成量が増加しました。本来Uchl1を発現しないヒトの細胞でも、強制的にセンスUchl1とアンチセンスUchl1を導入すると、UCHl1タンパク質の合成量が増加しました(図2)。また、タンパク質の合成量増加にはSINEB2が必要で、特に、アンチセンスUchl1内に存在する位置が重要であることが分かりました。
FANTOM3で同定されたアンチセンスRNAを使って、別の遺伝子(Uxt)でもSINEB2のタンパク質合成促進機能が確認され、その普遍性が示唆されました。
次にアンチセンスUchl1のタンパク質合成を促進する経路を調べるために、一般的なmRNAの翻訳開始機構であるCAP構造※8を抑制し、通常の翻訳機能を阻害しました。その結果、アンチセンスUchl1のSINEB2が、通常の翻訳とは異なる機能でmRNAのリボソームへの移行し、タンパク質合成を促進しました。また、タンパク質の生合成を制御する酵素mTORC1の阻害を阻害すると、通常は核に多く存在するアンチセンスUchl1を細胞質へ移行させることも分かりました。
このようなアンチセンスUchl1に見られる機能は、ストレスなどの外的要因により通常の翻訳開始機構を担う使う遺伝子発現が阻止された時に、必要なタンパク質を合成して生き残るための保存的機能である可能性があります。
以上から、mRNAのタンパク質翻訳を促進するには、「タンパク質の合成促進機能を持つSINEB2」と「目的のmRNAに特異的に結合する領域」の2つが必要と分かりました。
今後の期待
今回、mRNAの翻訳を制御するというncRNAに新たな機能を初めて見いだしました。タンパク質翻訳に関わる2つの領域を持つ配列をデザインすることで、目的とするタンパク質をコードするmRNAから効率的にタンパク質を合成することが可能です。今後、基礎研究用だけでなく医療などの実用的なタンパク質合成技術となる可能性が考えられます。また、ncRNAの新たな実用性のある機能分野を示したことで、未知なるncRNAの機能の可能性も期待できます。
原論文情報
- Claudia Carrieri, Marta Biagioli, Laura Cimatti, Anne Beugnet, Isidre Ferrer, Silvia Zucchelli, Stefano Biffo, Allistar Forrest, Piero Carninci, Elia Stupka, and Stefano Gustincich.
"Long non-coding antisense RNA controls Uchl1 translation through an embedded SINEB2 repeat". Nature,2012,doi: 10.1038/nature11508
発表者
理化学研究所
オミックス基盤研究領域 LSA要素技術開発グループ ゲノム機能研究チーム
チームリーダー ピエロ カルニンチ
お問い合わせ先
横浜研究推進部 企画課
Tel: 045-503-9117 / Fax: 045-503-9113
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.アンチセンスRNA
mRNAなど、ある機能を持つRNA(センスRNA)と相補的な塩基配列を持つ短いRNAで、センスRNAと2本鎖を形成することでそのセンスRNAが担うべきタンパク質の合成を阻害すると考えられる。 - 2.メッセンジャーRNA(mRNA)
タンパク質を翻訳できる塩基配列情報と構造を持ったRNA。mRNAはDNAから写し取られた遺伝情報に従い、タンパク質を合成(翻訳)する。 - 3.リボゾーム
すべての生物の細胞に存在するタンパク質合成を担う複合体。mRNAの遺伝情報を読み取ってタンパク質への翻訳が行われる場所である大小2つのサブユニットから成り、リボソームタンパク質とリボソームRNAから構成される。 - 4.FANTOM
理化学研究所のマウスゲノム百科事典プロジェクトで収集された完全長cDNAのアノテーション(機能注釈)を行うことを目的に、2000年に、理研ゲノム科学総合研究センター 遺伝子構造機能研究グループ(現・オミックス基盤研究領域)が中心となって結成した。哺乳動物(マウス)の遺伝子を網羅的に機能注釈することを主眼とする国際的研究コンソーシアム共同集団(Functional ANnoTation Of Mammalian genome)の略称。活動範囲は拡大され、2009年には遺伝子ネットワークの解明に成功。現在は様々な種類の細胞の遺伝子ネットワーク解析に取り組んでいる。現時点で、18カ国100機関以上が参加している。 - 5.ノンコーディングRNA(ncRNA)
DNAから転写されるが、タンパク質には翻訳されないRNAの総称。タンパク質翻訳以外の転写制御などの機能を担っていると考えられている。 - 6.理研ベンチャー
理研の研究成果を積極的に技術移転する方策の一環として、理研が経済社会の発展、科学技術の振興などの観点から有意義である企業を「理研ベンチャー」として認定し、支援する制度。理研は、知的財産権の実施許諾における優遇措置、共同研究実施に伴う施設利用に関する優遇措置などを行うことにより、理研ベンチャーに対する支援を行っている。 - 7.短鎖散在反復配列(SINE)
ゲノムの特定の塩基配列がコピーされ、これが再びゲノムに挿入されたもの。生物進化の過程において、ある生物のゲノムの特定の場所にSINEが挿入されると、これが子孫に受け継がれる。このことから、多数の生物のSINEを分析すると系統関係が分かる。最も良く見られるSINEは、Alu配列 (Alu sequence)。Alu配列は、約300塩基対の長さがあり、タンパク質コード配列を全く含まず、制限酵素AluIで認識されるため、“Alu”と命名された。SINEは、ヒトゲノムの13.5%を占め、そのうち約11%をAlu配列が占める。SINEは、その目的が判明しておらず、「利己的なDNA」とか「ジャンクDNA」とも呼ばれている。 - 8.CAP構造
真核生物のmRNAの5’末端にはCAP(キャップ)と呼ばれる特徴的構造が存在する。この構造は、転写後早い段階で酵素的な修飾により形成され、5’末端を保存するための構造と考えられる。また、翻訳開始因子は、CAP構造を目印にしてmRNAの先端に結合する。
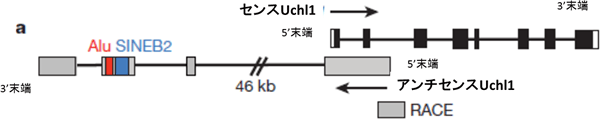
図1 センスUchl1とアンチセンスUchl1のゲノム上の配列構造
センスUchl1の黒い四角はエクソン(RNAに転写される部分)、白い四角は非翻訳領域を表す。アンチセンスUchl1の灰色の四角はエクソン。Alu、SINEB2は、反復配列領域。SINEB2と5’末端が重要であることが分かった。
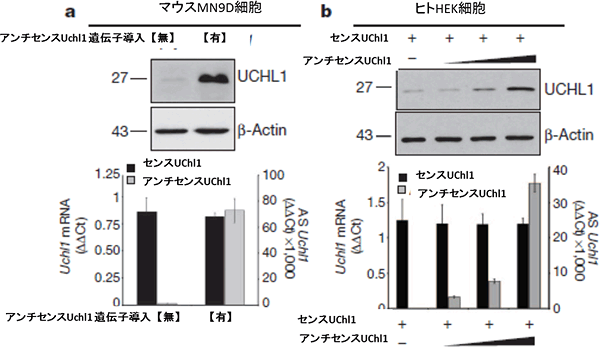
図2 マウス由来細胞株(MN9D)とヒト由来細胞株(HEK)でのUCHL1タンパク質発現
- (a) マウス由来細胞(MN9D)にアンチセンスUchl1を導入し、UCHL1タンパク質が発現した。βactinは比較対象。
- (b) ヒト由来細胞株(HEK)にセンスUchl1およびアンチセンスUchl1を導入し、UCHL1タンパク質が発現した。