ポイント
- Cmr複合体は6種類のタンパク質とRNAから成るらせん状の巨大な分子複合体
- Cmr複合体はウイルスのRNAを5カ所で分解し、ウイルスを破壊
- 高等動物が持つ複雑な獲得免疫システムの起源とその進化の解明に寄与
要旨
理化学研究所(理研、野依良治理事長)は、細菌が持つ免疫システムを担う巨大な分子複合体の「Cmr複合体」の構造と機能を解明しました。この発見は、高等動物が進化の過程で得た獲得免疫の起源とその進化を解明するために重要な知見となります。これは、放射光科学総合研究センター(石川哲也センター長)ビームライン基盤研究部の新海暁男先任研究員(現 横山構造生物学研究室 先任研究員)、上利佳弘特別研究員、同センター生物試料基盤グループの眞木さおり研究員、同センター米倉生体機構研究室の米倉功治准主任研究員、オランダのワーゲニンゲン大学のレイモンド スタールズ(Raymond H.J. Stalls)研究員、ジョン ファン デア オウスト(John van der Oost)教授らによる共同研究グループの成果です。
全ての生物は、外界の毒物や病原菌から生体を守るために免疫システムを備えています。免疫システムは、生物種に応じてさまざまな種類のシステムが存在し、生物の進化と同様に免疫システムも進化してきたと考えられています。免疫システムのなかでも獲得免疫は、ウイルスなどの抗原情報を記憶するなど高度な仕組みを持っていますが、このシステムの起源や進化の過程は、いまだ明らかになっていません。
細菌は、生命の歴史のなかでも初期に誕生した生物です。多くの細菌は、獲得免疫に似た「CRISPR-Cas(クリスパー-キャス)システム[1]」と呼ばれる防御システムを備えています。共同研究グループは、細菌のうちの一種である「高度好熱菌Thermus thermophilus HB8株[2]」をモデルとして、このシステムの全容を解明し、獲得免疫の起源や進化の一端を明らかにすることに挑みました。
共同研究グループは、CRISPR-Casシステムを担う6種のCasタンパク質[3]とRNA断片で構成されているCmr複合体を単離精製し、結合しているRNAの配列解析、標的核酸の切断活性や電子顕微鏡観察などによって詳細な構造と機能を解明しました。その結果、Cmr複合体はユニークならせん構造をしていることに加え、そのRNA断片は感染したウイルスのRNAの一部に結合することでウイルスを捕らえ、切断することでウイルスを破壊し感染を防ぐことが分かりました。
生物は古来より、ウイルスとの戦いを繰り返してきました。本研究は、生物が長い歴史の中で培ってきたウイルスに対する免疫・防御システムの原型や進化を知る上で重要な発見といえます。
本研究成果は、日本学術振興会の科学研究費補助金(課題番号:25440013)の助成を受けて行われ、米国の科学雑誌『Molecular Cell』オンライン版(10月10日付け:日本時間10月11日)に掲載されます。
背景
全ての生物は、外界の毒物や病原菌から生体を守るために免疫システムを備えており、生物種に応じてさまざまな種類が存在します。生物の進化と同様に、免疫システムも突然変異と環境適応を繰り返しながら進化してきたと考えられています。免疫システムは、自然免疫と獲得免疫の2つに大別されます。獲得免疫は、外界から入り込んだ病原菌(抗原)の一部を免疫システムに関わる分子(細胞)が取り込み、抗体を作って抗原を攻撃します。特に重要な機能の1つとして、その情報を記憶し、同じ抗原に再度感染したときも迅速に攻撃することがあります。この獲得免疫システムの起源やその進化の過程については、いまだ不明なことが多く明らかになっていません。
細菌は、生命の歴史のなかでも初期に誕生した生物です。多くの細菌は、獲得免疫に似た「CRISPR-Casシステム」と呼ばれる防御システムを持っています。CRISPR-Casシステムは、DNA領域「CRISPR領域(図1A)」と、その領域近傍のcas遺伝子群にコードされる「Casタンパク質群(図1B)」から構成されています。このシステムを持つ細菌は、ウイルスなどが侵入すると、ウイルスのDNAの一部を切り取り、細菌のゲノム上のCRISPR領域に組み込みます。ウイルスのDNAを組みこんだCRISPRのDNA配列は、RNAに変換され、複数の断片に切断されます。切断されたRNAをCRISPR RNA(crRNA)[4]と呼び、これらは、防御システムに関わるCasタンパク質と複合体を形成します。この細菌が、再度同じウイルスに感染しても、ウイルスのDNAやRNAと適合するcrRNAを持ったCasタンパク質複合体が反応し、そのウイルスを攻撃し排除することができます(図2)。
CRISPR-Casシステムは、同じ生物種の中でもCasタンパク質の種類やDNA/RNAを切断するメカニズムによって3つに分類され(タイプI、II、III)、さらに、幾つかのサブタイプに複雑に分かれています。これらの機能や構造を詳細に解析することで、生物の持つウイルスに対する免疫・防御システムの原型や進化が明らかになると考えられています。
共同研究グループは、高度好熱菌「Thermus thermophilus HB8株(図3)」をモデル生物として、各種CRISPR-Casシステムの全容を解明し、獲得免疫システムの原型や進化を探ることに挑みました。
研究手法と成果
T. thermophilusHB8株は、タイプI-E(Cascade)複合体[5]、タイプIII-A(Csm)複合体[6]、およびタイプ III-B(Cmr)複合体の3つの複合体を持ちます。共同研究グループは、T. thermophilusHB8株から、6種類のCasタンパク質(Cmr1, -2, -3, -4, -5, -6)とcrRNAから構成されるCmr複合体を単離・精製しました。次に、Cmr複合体を構成するcrRNAの配列を解析した結果、特定のCRISPR領域に由来するcrRNAが多く見つかりました(図3)。このことから、crRNAは無作為に結合されるのではなく、選択性があることが分かりました。さらに、Cmr複合体は、crRNAの配列に対して相補的な配列を含む一本鎖RNAを6塩基間隔で5カ所、切断することも分かりました(図4)。
また、Cmr複合体を電子顕微鏡により構造を観察した結果、その全体はユニークならせん状の構造をしていることが分かりました(図5A)。さらに、マススペクトル解析[7]で突き止めた Cmr複合体を構成するCasタンパク質とcrRNAとの分子量比の結果と、電子顕微鏡からの構造を総合的に解析したところ、各Casタンパク質の配置を推定できました(図5B)。それによると4つあるCmr4と呼ばれるCasタンパク質が約25 Å(オングストローム:1 Å は10-8 cm)間隔でらせん状に位置し、その間隔はRNAの6~7塩基間隔に相当していました。
Cmr複合体によってRNAが5カ所で切断されることから、中央部に位置している4つのCmr4ともう1つのCasタンパク質で切断されることが強く示唆されました。また、今回解明されたCmr複合体の全体構造は、既に報告されていた大腸菌由来のタイプI -E(Cascade)複合体と類似していたため(図5C)、これらの巨大な分子複合体は共通祖先から派生したものと考えられます。
以上の結果、T. thermophilus HB8株のCmr複合体は、感染したウイルスが持つmRNAの一部にcrRNAが結合することでCmr複合体中のCmr4らが5カ所で切断し、ウイルスを破壊して感染を防ぐと考えられます(図6)。
今後の期待
T. thermophilusHB8株由来のタンパク質は安定性が高く、構造解析に適しているので、大型放射光施設SPring-8[8]やSACLA[9]を用い、超分子複合体の高分解能の構造や分子の動的特性を解析し、各Casタンパク質の役割や作用メカニズムをより詳しく解析することが可能と考えています。また、T. thermophilus HB8株が持つ他のサブタイプ複合体の構造と機能を同様に解析し、1つの細胞が持つCRISPR-Casシステムを体系的に理解することによって、獲得免疫システムの起源、進化に対する新たな知見が得られると期待できます。
原論文情報
- Staals, R.H.J.,* Agari, Y.,* Maki-Yonekura, S.,* Zhu, Y., Taylor, D.W., van Duijn, E., Barendregt, A., Vlot, M., Koehorst, J.J., Sakamoto, K., Masuda, A., Dohmae, N., Schaap, P.J., Doudna, J.A., Heck, A.J.R., Yonekura, K., van der Oost, J.,† and Shinkai, A.† "Structure and activity of an RNA-targeting Type III-B CRISPR-Cas complex in Thermus thermophilus", Molecular Cell, 2013. doi:10.1016/j.molcel.2013.09.013
(*equal contribution, †corresponding author)
発表者
理化学研究所
上席研究員研究室 横山構造生物学研究室
先任研究員 新海 暁男 (しんかい あけお)
(当時 放射光科学総合研究センター ビームライン基盤研究部 先任研究員)
お問い合わせ先
放射光科学研究推進室
Tel: 0791-58-0900 / Fax: 0791-58-0800
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.CRISPR-Casシステム
細菌が持つ、ウイルス(ファージ)やプラスミドなどの侵入核酸に対する免疫・防御システム。CRISPR領域は、約25~50 bpの回文様の配列が約25~50 bpの間隔(スペーサー)で繰り返し存在するDNA領域と、CRISPR近傍の遺伝子群にコードされているCasタンパク質群とから構成されている。 - 2.高度好熱菌 Thermus thermophilus HB8株
静岡県伊豆半島にある峰温泉から発見された、85℃という極限環境で生育できる細菌。熱水中で生きている細菌(好熱菌)は全生物の共通祖先に近い位置にあり、原始生命の基本的特徴が凝縮されていると考えられている。11箇所のCRISPR領域と約30種類の cas遺伝子を持つ。 - 3.Casタンパク質
CRISPR領域の近傍に位置している cas遺伝子にコードされているタンパク質で、これまで45種類のファミリーが見つかっている。また、数種類のサブユニットから成る巨大な分子複合体を形成しているものもある。これらのタンパク質は、CRISPR-Casによる免疫・防御システムに関与している。 - 4.CRISPR RNA(crRNA)
crRNAは、Casタンパク質に結合して複合体を形成する。このCasタンパク質-crRNA複合体は、侵入DNAあるいはRNAの一部分に相補的に結合し、分解する。 - 5.タイプI-E(Cascade)複合体
Cse1、Cse2、Cas7、Cas5e、Cas6eタンパク質とcrRNAが、1:2:6:1:1:1分子で形成された巨大な分子複合体(405 kDa)。Cascade複合体は、pre-crRNAを分断してcrRNAを生成し、複合体に取り込む。さらに、Cas3タンパク質と結合し、標的DNAを分解する。大腸菌由来のCascade複合体の研究が進んでおり、詳細な機能メカニズムや電子顕微鏡構造が明らかにされている。 - 6.タイプIII-A(Csm)複合体
5種類(Csm1, -2, -3, -4, -5)のタンパク質サブユニットとcrRNAとから成る複合体で、DNAの切断に関与している。 - 7.マススペクトル
高電圧をかけた真空中で試料をイオン化すると、静電力によって試料は装置内を飛行する。飛行しているイオンを電気的・磁気的な作用等により質量電荷比に応じて分離し、その後それぞれを検出することで、質量電荷比を横軸、検出強度を縦軸とするマススペクトルを得ることができる。このスペクトルから試料の分子量を求めることができる。 - 8.大型放射光施設SPring-8
理研が所有する、兵庫県の播磨科学公園都市にある世界最高性能の大型放射光施設。SPring-8の名前はSuper Photon ring-8GeVに由来する。放射光(シンクロトロン放射)とは、電子を光とほぼ等しい速度まで加速し、電磁石によって進行方向を曲げたときに発生する、細く強力な電磁波のことである。SPring-8では、遠赤外から可視光線、軟X線を経て硬X線に至る幅広い波長域で放射光を得ることができるため、原子核の研究からナノテクノロジー、バイオテクノロジー、産業利用や科学捜査まで幅広い研究が行われている。 - 9.SACLA
理研と高輝度光科学研究センターが共同で建設した日本で初めてのXFEL施設。科学技術基本計画における5つの国家基幹技術の1つとして位置付けられ、2006年度から5年間の計画で建設・整備を進めた。2011年3月に施設が完成し、SPring-8 Angstrom Compact free electron LAser の頭文字を取ってSACLAと命名された。2011年6月に最初のX線レーザーを発振、2012年3月から共用運転が開始され、利用実験が始まっている。大きさが諸外国の同様の施設と比べて数分の1と、コンパクトであるにも関わらず、0.1ナノメートル以下という世界最短波長のレーザーの生成能力を有する。
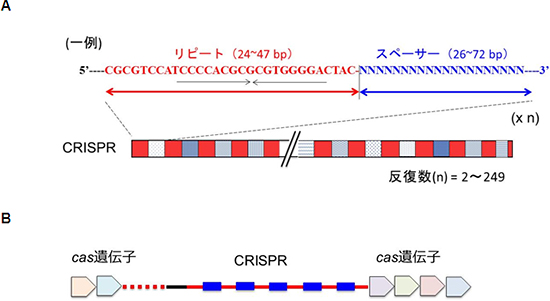
図1 CRISPR領域とcas遺伝子
- A.CRISPR領域は、約25~50 bpの回文様のリピート配列(図中赤色部分)が、25~50 bpの間隔(スペーサー配列)(図中青色部分)を介して反復している領域。スペーサーの配列には共通性はない。反復数は多いもので249回に及ぶ。
- B.cas遺伝子はCRISPR領域の近傍に存在する。
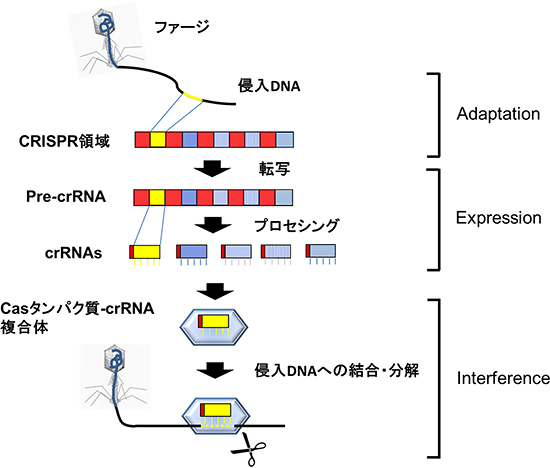
図2 CRISPR-Casシステムの作用モデル(例)
CRISPR-Casシステムには、主に3つの段階がある。ウイルス(ファージ)由来のDNAなど、細胞へ侵入したDNAの一部が切り取られ、新たなスペーサーとしてCRISPR領域に挿入される(Adaptation phase)。CRISPR領域は転写されてpre-crRNAが生じ、pre-crRNAは切断されてcrRNAが生ずる(Expression phase)。crRNAはCasタンパク質と共に複合体を形成し、侵入DNA由来のcrRNAを持つ複合体は侵入DNAに相補的に結合し、分解する(Interference phase)。
CRISPR領域の数や配列、cas遺伝子の種類は生物種毎に異なり、侵入DNAから転写されたRNAを分解するCRISPR-Casタイプもある。
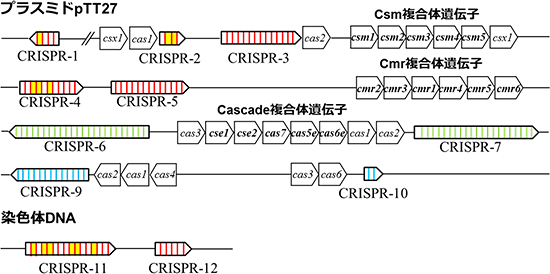
図3 Thermus thermophilus HB8株の遺伝子に存在するCRISPR領域とcas遺伝子
CRISPR領域は、プラスミドpTT27上に9箇所、染色体DNA上に2箇所存在する(CRISPR-8は除外した)。リピートの塩基配列によって3種類(図中赤色、緑色、青色部分)に分類できる。タイプIII-A(Csm)複合体遺伝子、タイプIII-B(Cmr)複合体遺伝子、タイプI-E(Cascade)複合体遺伝子を太字で示した。
Cmr複合体のcrRNAの配列を解析した結果、CRISPR-1、-2、-4、-11に存在する特定のスペーサー由来のcrRNA(図中黄色部分)が多く見つかった。このことから、crRNAは無作為に結合されるのではなく、選択性があることが分かった。
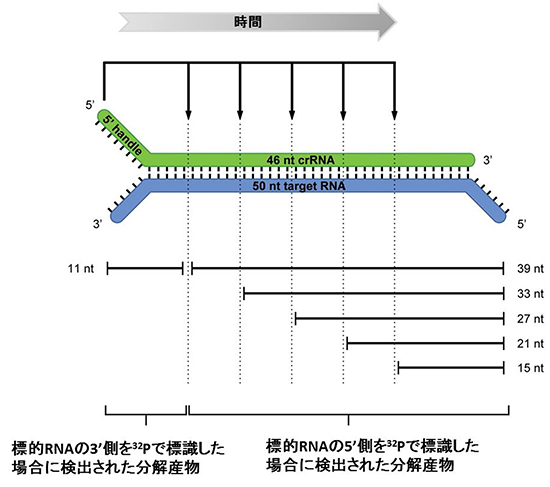
図4 Thermus thermophilus のCmr複合体による標的RNAの切断様式
Cmr複合体は、複合体が持つcrRNA(46 nt crRNA)と相補的な配列を含む標的RNA(50 nt target RNA)と結合する。結合後、標的RNAはその3’側から順に6塩基間隔で5箇所、切断される。
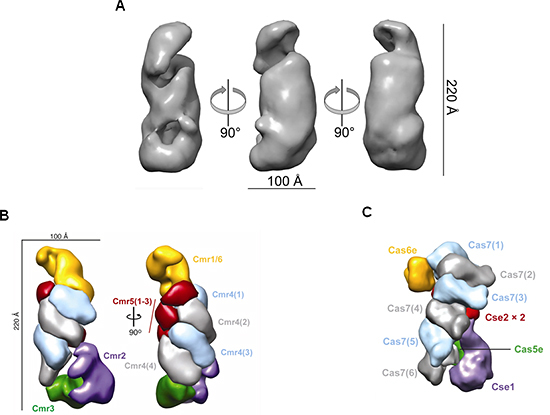
図5 Cmr複合体の電子顕微鏡による構造
- A.Thermus thermophilus Cmr複合体の電子顕微鏡構造。
- B.マススペクトル解析、電子顕微鏡構造などから推定したCmr複合体のCasタンパク質の構成。分子量比は1:1:1:4:3:1の割合で、4分子のCmr4タンパク質と3分子のCmr5タンパク質がらせん状に位置している。
- C.大腸菌由来タイプI-E(Cascade)複合体の電子顕微鏡構造から推定されたCasタンパク質の構成。6分子のCas7タンパク質がらせん状に位置している。
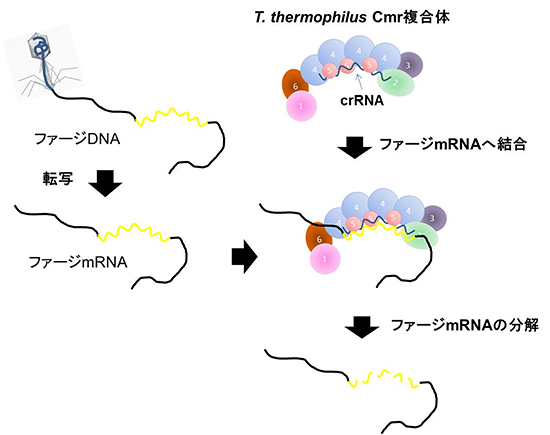
図6 Thermus thermophilus Cmr複合体の作用メカニズム
Cmr複合体は、細菌に感染したウイルス(ファージ)由来のmRNAのうち、Cmr複合体のcrRNAと相補的な配列を持った部分に結合し、ファージのmRNA を5箇所で切断する。Cmr複合体の中央部に位置している4つのCmr4ともう一箇所別のCmrが分解活性の中心と考えられる。