ポイント
- 脊椎動物の発生過程では、転写開始点を選択するメカニズムが大きく切り替わる
- 1つのプロモーターに対して、2つの転写制御のメカニズムが使い分けられる
- ゲノム上の遺伝子発現制御をより詳細に理解する新しい枠組みを提示
要旨
理化学研究所(理研、野依良治理事長)は、小型魚類のゼブラフィッシュの初期胚の解析により、遺伝子の転写開始点が2つの異なるメカニズムによって決定され、遺伝子発現機能を持つ塩基配列(プロモーター配列[1])1つに対して2つのメカニズムが独立に作用する新しい転写制御の仕組みを発見しました。これは、理研ライフサイエンス技術基盤研究センター(渡辺恭良センター長)機能性ゲノム解析部門のピエロ・カルニンチ部門長、ゲノミクス微量技術開発ユニットのシャルル・プレシ ユニットリーダー、トランスクリプトーム研究チームのアナ・マリア・スズキ技師らと、ノルウェー・ベルゲン大学、英国・インペリアル カレッジ ロンドンおよびバーミンガム大学、オランダ・エラスムス医療センター、ドイツ・カールスルーエ大学との国際共同研究グループによる成果です。
ゲノムDNAの塩基配列がRNAに転写される際、最初に写し取られる塩基をゲノムDNA上の転写開始点と呼びます。転写開始点の近傍は、遺伝子発現のタイミングや発現量の制御に重要な役割をもつ領域です。これまでの研究から、プロモーターの配列と転写開始点の関係では2つのタイプの存在が明らかになっていましたが、転写開始点を決定する仕組みについてはよく分かっていませんでした。
共同研究グループは、脊椎動物の発生初期において、卵形成時に蓄えられた卵由来のRNAがなくなり、受精卵ゲノムから新たなRNAの転写が始まる現象に着目しました。理研は、特定の細胞や発生中の個体で発現している遺伝子の転写開始点を、網羅的かつ1塩基レベルの高い精度で解読する遺伝子解析技術「CAGE法[2]」を開発しています。これを用いてゼブラフィッシュの初期胚における遺伝子の転写開始点を網羅的に調べた結果、発生の進行に伴い転写開始点を決定するメカニズムが切り替わることを発見しました。詳しい解析により、卵由来のゲノムの発現には転写開始点の上流の領域が関わり、受精卵ゲノムの発現には転写開始点の下流の領域とクロマチン構造[3]の変化が関わっていることが分かりました。さらに、1つのプロモーターが両方の領域の特徴を併せ持ち、見かけ上は恒常的な遺伝子発現が起きている場合でも、2つの転写機構の制御をそれぞれ独立に受けて制御を独立に受けて転写開始点が決定されていました。これらの結果は、遺伝子発現制御メカニズムの切り替えが、遺伝子を卵と胚で発現させるための普遍的な仕組みであることを示唆しています。
ゲノム上の制御領域には、遺伝子発現に関わるさまざまな情報を受け取る配列が重層的に埋め込まれており、その機能の高精度な配列解析が待たれていました。プロモーターの配列を読む「読み方」が発生の過程で使い分けられていることを明らかにした今回の発見は、複雑だった遺伝子制御機構の解釈をシンプルにします。
本成果は、英国の科学雑誌『Nature』(2月16日付け : 日本時間2月17日)に掲載されました。
背景
細胞は状況に応じて、発現する遺伝子の種類や量を変化させます。遺伝子発現はさまざまな細胞内外のシグナルの制御を受けますが、最終的にはプロモーターに基本転写因子が結合し、RNAポリメラーゼが転写開始点からRNAの合成を始めることで遺伝子の転写が開始されます。従って、細胞で発現している遺伝子の転写開始点をそれぞれ網羅的に解析し、それらのプロモーターを比較した結果は、その細胞ではどのようなルールでゲノムDNAの配列が読まれ、どの遺伝子の発現が制御されているかを特定する重要な手がかりとなります。
理研が開発した遺伝子解析技術の1つであるCAGE法は、少量のRNA試料からでも末端配列を決定でき、転写開始点を網羅的かつ1塩基レベルの正確さで解読できる方法です。これまでにヒト由来の細胞株などをCAGE法で解析した結果、プロモーターには、1つの転写開始点を決めるものと、複数の転写開始点を散在させるものの2つのタイプがあることが分かっています。しかし、どのように使い分けているかは不明でした。
動物の発生過程では、遺伝子発現の制御が大きく変わる時期があります。卵が精子と受精した後しばらくの間は、発生は母体内での卵形成時に蓄えられた卵由来のmRNAやタンパク質の働きで進行しますが、やがて受精卵(胚)に由来するゲノムDNAからの転写が始まります。小型魚類のゼブラッフィッシュの発生では、この切り替わりは受精卵が10回目の細胞分裂を経た時期にあたり、MBT(中期胞胚遷移)[4]と呼ばれます。今回の研究では、MBTの前後で転写されるゲノムDNAが切り替わる際、それぞれの転写開始点のパターンがどのように変化するかをCAGE法で解析し、さらに転写開始に影響を与えるとされるクロマチン構造についても調べました。
研究手法と成果
共同研究グループは、未受精卵からMBTを経て器官形成期にいたる12の発生段階のゼブラフィッシュ胚を用い、CAGE法でそれぞれの時期に発現する遺伝子の転写開始点を決定しました。その結果、1つのプロモーターからの遺伝子発現であっても、卵ゲノム由来の転写開始点と胚ゲノム由来の転写開始点が異なり、MBT前後で切り替わる例が多数見つかりました(図1)。また、全ての転写産物の転写量変化を調べた結果、特にMBT前後で大きく変化するものが多いことが分かりました。これは、ゼブラフィッシュの細胞内の全RNA、つまりトランスクリプトーム[5]がMBTを境に大きく変化することを示しています。
転写開始点が切り替わるプロモーターの配列を詳しく調べると、卵ゲノム由来の転写開始点の上流30bp付近にはチミン(T)とアデニン(A)が連続して並ぶ配列(Wボックス・モチーフ[6])がありました(図2)。このDNA配列は転写開始を決定する機能があることが一般的に知られており、これが卵由来ゲノムに特徴的な転写開始点の選択に関わっていることが予想されました。実際に、プロモーターからWボックスを欠損させても、胚ゲノムの転写には影響はなく、卵由来ゲノムの転写開始点のみが消失しました。一方、胚ゲノム由来の転写開始点の上流にはWボックスはなく、下流50bp付近に、グアニン(G)とシトシン(C)が連続して並ぶ配列からチミンとアデニンが連続する配列に変わる境界(SS│WW境界[7])が存在しました。クロマチン構造の解析から、このSS│WW境界の直下流領域には、転写開始を促進する特徴的なヌクレオソーム構造[8]がMBT後に形成されることが分かりました(図3)。これらの結果は、卵ゲノムと胚ゲノムの転写開始はそれぞれ異なるメカニズムの制御を受けており、MBTの前と後で1つのプロモーターから異なる転写開始点が選択されるのは、2つのメカニズムがそれぞれ独立に1つのプロモーターに作用している結果であることを示します。つまり遺伝子発現制御メカニズムの切り替えは、MBT前後で発現を変化させる遺伝子だけではなく、見かけ上は恒常的に発現している遺伝子にも適用される、卵と胚での遺伝子発現を制御する普遍的な仕組みであることが示唆されます。
今後の期待
プロモーターの役割を正確に捉えるのが困難である理由の一つは、短いDNA配列の中に複雑な遺伝子発現パターンを決める情報が重層的に埋め込まれているためです。今回の発見は、プロモーター配列の読み方を使い分ける2つの「文法」が存在することを示すものであり、プロモーターの構造と機能を解読する新たな枠組みとなると期待できます。また、発生過程におけるMBTの前と後のように、分化した細胞と未分化な細胞では転写制御メカニズムが大きく異なる可能性があります。今回の手法を幹細胞やiPS細胞(人工多能性幹細胞)に応用し、未分化状態から目的とする細胞への分化過程に関わる転写開始の仕組みの解明や、転写制御メカニズムの操作による細胞の分化制御に利用できると期待できます。
これまで詳細が分からなかった転写開始点の同定を可能にするCAGE法は、細胞の個性を決定する転写制御ネットワークを解明する強力なライフサイエンス技術であり、今後も生命科学・医学の各分野での応用を進めていきます。
原論文情報
- Vanja Haberle, Nan Li, Yavor Hadzhiev, Charles Plessy, Christopher Previti, Chirag Nepal, Jochen Gehrig, Xianjun Dong, Altuna Akalin, Ana Maria Suzuki, Wilfred van IJcken, Olivier Armant, Marco Ferg, Uwe Strähle, Piero Carninci, Ferenc Müller & Boris Lenhard. “Two independent transcription initiation codes overlap on vertebrate core promoters”. Nature, 2014 (doi: 10.1038/nature12974)
発表者
理化学研究所
ライフサイエンス技術基盤研究センター 機能性ゲノム解析部門
部門長 CARNINCI Piero(ピエロ・ カルニンチ)
技師 香月 美穂(かつき みほ)
お問い合わせ先
独立行政法人理化学研究所
ライフサイエンス技術基盤研究センター
チーフ・サイエンスコミュニケーター 山岸 敦(やまぎし あつし)
Tel: 078-304-7138 / Fax: 078-304-7112
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.プロモーター配列
遺伝情報は、A(アデニン)、T(チミン)、G(グアニン)、C(シトシン)という4種類の塩基の配列によってゲノムDNAに書かれている。プロモーター配列とは遺伝子を発現させる機能を持つ塩基配列のこと。プロモーターがないと遺伝子は発現しない。 - 2.CAGE法
Cap Analysis of Gene Expressionの略。理研が独自に開発した方法で、耐熱性逆転写酵素やcap捕捉法を組み合わせて転写物の5’末端の塩基配列を決定する実験手法。この塩基配列を読み取ってゲノム配列と照らし合わせて、どこから転写が始まっているかを調べることができる。遺伝子の転写開始点をゲノムワイドに同定できる。 - 3.クロマチン構造
真核生物のゲノムDNAは、ヒストンなどのタンパク質と結合し、高度に凝縮した状態で存在する。この構造をクロマチン構造と呼ぶ。局所的なクロマチン構造の変化により、転写因子などの染色体DNAへの接近しやすさを制御する。 - 4.MBT(中期胞胚遷移)
mid-blastula transitionの略。胚発生の過程で、胞胚期に卵形成時に蓄えられた卵ゲノム由来のRNAが消失し、受精卵のゲノム(胚ゲノム)からの転写活性が高まる過程で起きる現象の総称。 - 5.トランスクリプトーム
細胞内の全DNAの塩基配列情報をさす「ゲノム」に対し、細胞内の全転写産物(全RNA)をトランスクリプトームと呼ぶ。その大半はタンパク質の情報を持たないノンコーディングRNA (ncRNA) であり、エピジェネティック制御やタンパク質合成制御、幹細胞性の制御などさまざまな機能に関わっている。 - 6.Wボックス・モチーフ
プロモーター配列のなかでも、TTGACC もしくは TTGACTの配列を持つもの。 - 7.SS│WW境界
2つの塩基WW (W = A or T)の連続が、SS(S = G or C)の連続へと切り替わる境界のこと。ヌクレオソーム構造の決定に関わることが知られている。 - 8.ヌクレオソーム構造
細胞核内では、約150塩基対(bp)のDNAが、ヒストン8量体の周りを1.65回巻き付いており、これをヌクレオソームと呼ぶ。電子顕微鏡でヌクレオソームを観察すると、細長い紐状のDNAがヒストンに巻きついた複数のヌクレオソーム構造の存在が確認できる。
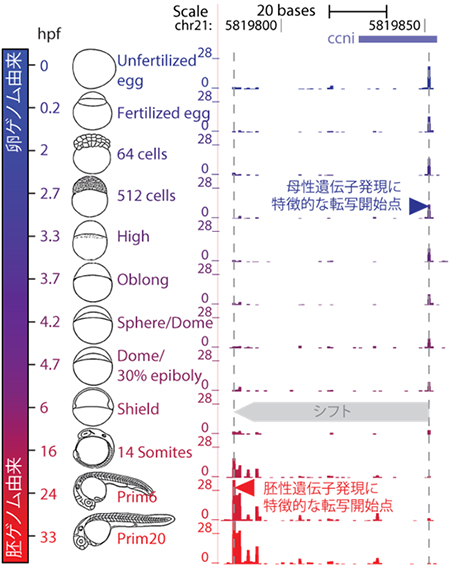
図1 MBT前後の転写開始点
ゼブラフィッシュの発生段階(左)と、MTB(中期胞胚遷移)前後の転写開始点の位置(右)。ここでは、卵ゲノム由来の転写開始点(青色)と胚ゲノム由来の転写開始点(赤色)が異なる例として、サイクリン1遺伝子(ccni)のプロモーター領域を示した。受精後6時間後(6hpf)のシールド期において、転写開始点の切り替わりが起きていることが分かる。
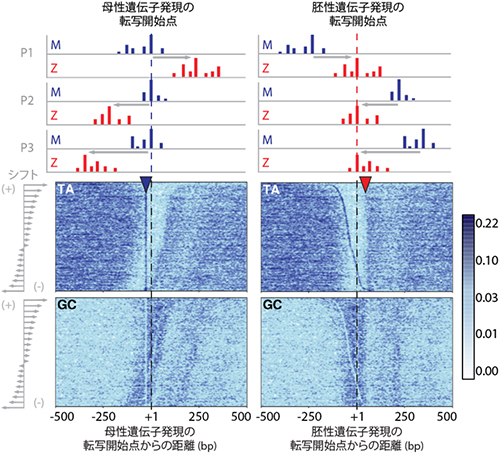
図2 転写開始点の選択に関わる上流配列と下流配列
転写開始点が切り替わる911個のプロモーターについて、卵ゲノム由来の転写開始点(青)もしくは胚ゲノム由来の転写開始点(赤)を基準にDNA配列を比較し、転写開始点の近傍に特徴的な配列を求めた。
- 上: 代表的な転写開始点の構造で、転写開始点シフトが下流に向かうもの(P1)と、上流に向かうもの(P2およびP3)について、CAGE法で得た最も頻度の高い転写開始点を+1として並べることを示す。
- 下: 上記ルールで並べた911個のプロモーター配列について、チミン(T)とアデニン(A)が連続で並ぶ領域とグアニン(G)とシトシン(C)が連続で並ぶ領域を可視化したもの。卵ゲノム由来の転写開始点の上流に30bp付近にTAの集中(Wボックス)が、胚ゲノム由来の転写開始点の下流50bp付近にTAとGCの境界(SS│WW境界)があることが分かる。
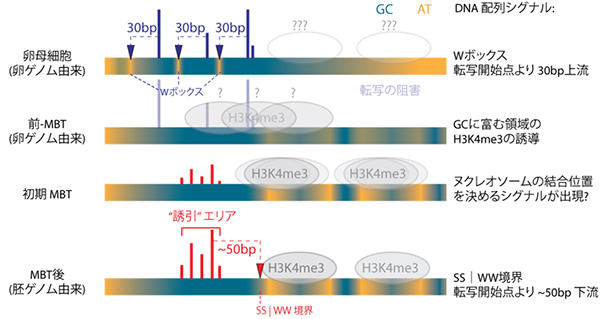
図3 MBT前後の転写開始メカニズムの変化
卵ゲノム由来の転写開始点(青)の30bp上流にはWボックスが存在し、また胚ゲノム由来の転写開始点(赤)の下流50bpにあるATとGCの境界(SS│WW境界)の直下に、転写開始を促進するヒストン(H3K4me3)が位置する。緑はGC富む領域、黄色はATに富む領域を示す。