2015年3月12日
理化学研究所
形態形成などに関わるHhシグナル伝達系の分子機構の一端を解明
-Sufu遺伝子はHhシグナル伝達系の最下流に位置する転写因子を制御-
要旨
理化学研究所(理研)バイオリソースセンター新規変異マウス研究開発チームの牧野茂 開発研究員と権藤洋一チームリーダーらの共同研究グループ※は、形態形成や発がんに重要な役割を果たす「ヘッジホグ(Hh)シグナル伝達系[1]」の、新たな活性制御機構の一端を明らかにしました。
Hhシグナル伝達系は、ほ乳類胎児の発生において神経、四肢などを含むさまざまな器官や臓器の形作りに重要な役割を担っています。また、成体ではHhシグナル伝達系の制御異常が発がんにつながることが知られています。Hhシグナル伝達系に関する研究は、胎児の体の形作りや成体での恒常性の維持に関する基本的メカニズムの解明に加え、病態の理解や治療法、創薬ターゲット開発にも役立つことから、多くの研究が行われています。しかし、Hhシグナル伝達系の分子レベルでの制御機構は極めて複雑であり、十分に解明されていませんでした。
これまでの研究により、Hhシグナル伝達系において最下流に位置する活性化型のGli転写因子(Gli1、Gli2)と抑制型のGli転写因子(Gli3)[2]が、Sufuタンパク質[3]によって制御されることが明らかにされています。特定の遺伝子の機能を調べるための最も標準的な手法は「遺伝子ノックアウト[4]」ですが、Sufu遺伝子は、ノックアウトすると胎生致死となり、発生後期や生後での遺伝子機能の解析ができません。また、ノックアウトでSufuタンパク質が完全になくなるため、活性化型と抑制型という反対の機能を持ったGli転写因子群のバランスを制御するSufuタンパク質の機能の解明も不可能でした。
共同研究グループは、遺伝子機能をノックアウトではなく一部の機能のみを変えて解析できるように、Sufu遺伝子の機能を部分的に変えた点突然変異マウスを16系統開発しました。次に、その中で重度の多指症(手足の形状異常)や神経形成不全を起こした1系統について、変異Sufuタンパク質の活性を、培養細胞や胚で解析しました。この系統は、Sufuタンパク質を構成するアミノ酸のうち、396番目のスレオニン残基に変異が生じていたため、抑制型のGli3の制御に影響をもたらしていましたが、活性化型のGli1、Gli2へは何ら影響がありませんでした。このことから、Sufuタンパク質の396番目のスレオニンが、Gli3のみを制御する残基として働いていることがはじめて明らかになりました。
今回開発した点突然変異マウスを利用することにより、神経や四肢の発生および発がんの基本的メカニズムの解明がさらに進むと期待できます。本研究成果は、米国のオンライン科学雑誌『PLOS ONE』(3月11日付け:日本時間3月12日)に掲載されます。
※共同研究グループ
理化学研究所
バイオリソースセンター 新規変異マウス研究開発チーム
開発研究員 牧野 茂(まきの しげる)
チームリーダー 権藤 洋一(ごんどう よういち)
石井分子遺伝学研究室
上席研究員 石井 俊輔(いしい しゅんすけ)
和歌山県立医科大学
研究員 松丸 大輔(まつまる だいすけ)
トロント小児病院(カナダ)
教授 シーシー・ホイ
背景
Hhシグナル伝達系(図1)は、ほ乳類胎児の発生において体の形作りに中心的な役割を果たすシグナル伝達系の1つとして、多くのグループによって研究が進められてきました。また、近年、ヒトのがんの25%でHhシグナル伝達系の制御異常が見られる注1)ことから、成体では、発がんにも関与することが明らかになっています。
これまでに、ノックアウトマウスを用いた遺伝学的な解析により、Hhシグナル伝達系に関わる多くの遺伝子が同定され、その伝達経路の骨組みが明らかにされてきました。Sufuタンパク質はその1つで、伝達経路の最下流に位置づけられるGli転写遺伝子群(Gli1、Gli2、Gli3)を直接制御することが明らかになってきました。しかし、Sufu遺伝子をはじめとして、Hhシグナル伝達系に関わる遺伝子の多くは必須遺伝子であるため、ノックアウトすると発生初期で胎生致死となり、発生後期や生後での遺伝子機能の解析ができませんでした。
また、Sufuタンパク質は、活性化型(Gli1、Gli2)と抑制型(Gli3)の両方のGli転写因子群を同時に、かつバランスを取りながら制御します。そのため、sufu遺伝子をノックアウトして形態異常などの特徴的な表現型を確認できても、活性化型と抑制型のどちらのGli転写因子の制御異常に基づくのか、どのようにバランスの破綻をもたらすのか、などを切り分けて解析できませんでした。
そこで、共同研究グループは、ノックアウトではなく遺伝子機能を部分的に変えた点突然変異マウスを開発し、発生後期以降のHhシグナル伝達系における、Sufuタンパク質によるGli転写因子の活性制御機構の解明を目指しました。
注1)Lum L and Beachy P. The Hedgehog response network: sensors, switches, and routers. Science. 304: 1755-9 (2004)
研究手法と成果
共同研究グループは、まず、マウスのゲノムDNAを1塩基レベルで機能解析するシステム注2)を利用し、「理研ENU誘発マウス点突然変異ライブラリー[5]」から、16系統のSufu点突然変異マウスを確立しました。その中の、7系統の変異マウスでは、Sufuタンパク質を構成するアミノ酸の配列に変化が生じていました(図2)。今回の解析には、重度の多指症(手足の形状異常)や神経形成不全を起こした1系統を使用しました。この系統は、Sufuタンパク質の396番目のスレオニン残基[6](T396)がイソロイシン残基[7]に変化したホモ接合体[8](以下、T396I変異マウス)です。
これまでの研究から、Sufu遺伝子を完全にノックアウトすると、四肢や頭部顔面の形態が形成される以前に死亡することが知られています。実際、Sufu遺伝子をノックアウトした場合と同じ特徴を示すR146X変異マウスは発生10日で死亡してしまいます(図3B)。一方、Sufu遺伝子の機能を部分的に変えたT396I変異マウスの胚を解析したところ、発生後期まで生き延び、形態形成も進みました(図3D、E)。これにより、T396I変異マウスを使えば、これまで不可能だった四肢や頭部形態形成期でのSufuタンパク質の機能解析が可能になりました。
また、このT396I変異マウスの頭部や四肢の形態は、Gli3遺伝子のみを完全に欠損した突然変異マウスによく似ていることから、T396Iタンパク質は、Gli3を制御する活性が低下する可能性が考えられました。そこで、T396I変異マウスの培養細胞や胚を詳細に調べたところ、変異したT396Iタンパク質はGli3を制御する機能を失っていること、一方、Gli1、Gli2を制御する機能はほぼ正常であることが明らかになりました。
これまでの研究で、Gli1とGli2はHhシグナル伝達系を活性化し、Gli3は抑制することが分かっています。以上の実験結果から、Hhシグナル伝達系において抑制型のGli3は、Sufuタンパク質の396番目のスレオニン残基に依存したシステムにより制御されていること、一方、活性化型のGli1、Gli2は、396番目のスレオニン残基に依存しないシステムにより制御されることが分かりました(図4)。
注2)2008年9月10日プレスリリース「1塩基レベルのゲノム機能解明が可能に」
今後の期待
T396I変異マウスを利用することで、Gli1/Gli2とGli3を制御する2つの異なった分子メカニズムの存在を示唆できました。今後、2つの分子メカニズムの制御に関わる因子を同定していくことで、Gli転写因子の活性機構の全貌を明らかにできると期待できます。
また、GliやGli活性制御に関わる遺伝子は、抗がん剤開発のターゲットとしても有望です。Hhシグナル伝達系のより下流の遺伝子やシグナルを標的とする方がより絞り込んだ発がん機構をターゲットとできるため、「副作用」の少ない創薬を期待できます。Sufu遺伝子の機能の解明は、疾患の原因解明や創薬開発に役立つ可能性があります。
T396I変異マウスは、理化学研究所バイオリソースセンターより入手可能で、「RBRC06556」という固有番号により検索できます。今後、このマウスを用いたさらなる成果が期待できます。
原論文情報
- Shigeru Makino, Olena Zhulyn, Rong Mo, Vijitha Puviindran, Xiaoyun Zhang, Takuya Murata, Ryutaro Fukumura, Yuichi Ishitsuka, Hayato Kotaki, Daisuke Matsumaru, Shunsuke Ishii, Chi-Chung Hui, Yoichi Gondo., "T396I Mutation of Mouse Sufu Reduces the Stability and Activity of Gli3 Repressor.", PLOS ONE, doi:10.1371/journal.pone.0119455
発表者
理化学研究所
バイオリソースセンター 新規変異マウス研究開発チーム
開発研究員 牧野 茂(まきの しげる)
チームリーダー 権藤 洋一(ごんどう よういち)
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.Hhシグナル伝達系
細胞外に分泌されたHhタンパク質から、受容体タンパク質やSufuタンパク質を含む複数のタンパク質を介して、最下流のGli転写因子群(Gli1、Gli2、Gli3)にシグナルが伝達される経路。Hh受容細胞では、Hhタンパク質の濃度に依存してGli転写因子の活性が変化する。そのため、発生期の四肢や神経管の細胞は、Hhタンパク質の濃度に依存した時空間的に正しい位置情報を得て、適切な細胞に分化し、体の形が作られる。 - 2.活性化型のGli転写因子(Gli1、Gli2)と抑制型のGli転写因子(Gli3)
Hhシグナル伝達系は、Sufuタンパク質を含めた複数のタンパク質を介して、最終的に3つのGli転写因子群に伝達される。構造的には3つとも類似したタンパク質でありながら、Gli1とGli2はHhシグナルの活性を促進し、Gli3は抑制する。Gli転写因子群は、細胞内の局在、安定性、核内での活性など、複数の段階で複雑に制御されると考えられているが、その全貌は明らかになっていない。 - 3.Sufuタンパク質
細胞膜と核との間でHhシグナルを仲介するタンパク質。既知のタンパク質との類似性がほとんど見られないことから、生化学的機能は不明。一方、既に作製されたノックアウトマウスの遺伝学的解析により、Hhシグナル伝達系に必須で、膜タンパク質Smoの下流かつGli転写因子群の上流で働くことが明らかになっている。 - 4.ノックアウトマウス
遺伝子操作により人為的に標的とする遺伝子を破壊した変異マウス。ノックアウトマウス個体に生じた変化を調べることで、破壊した遺伝子の機能を明らかにできる。 - 5.理研ENU誘発マウス点突然変異ライブラリー
ゲノムDNAの1塩基をさまざまな箇所で変化させた点突然変異マウス約1万匹分の凍結精子とゲノムDNAのアーカイブ。ノックアウトマウスとは異なり、1塩基レベルで、個体での遺伝子機能を解析できるシステム。点突然変異マウスは、理研バイオリソースセンターから広く研究者に提供されている。 - 6.スレオニン残基
アミノ酸の1種。本研究で開発・利用したマウスの場合、 Sufu遺伝子の396番目のスレオニン残基(T396)は、‘ACC’というゲノムDNA3文字遺伝暗号で書かれている。 - 7.イソロイシン残基
アミノ酸の1種。本研究で開発・利用したマウスの場合、上記スレオニン残基の遺伝暗号が‘ATC’に書き替わった結果、イソロイシン残基の遺伝暗号となった。スレオニン残基の遺伝暗号との違いはCとTのわずか1文字。 - 8.ホモ接合体
哺乳類は遺伝子セット(ゲノム)を母から1セット父から1セット受け継ぎ、計2セットもっている。 Sufu遺伝子も1つずつ両親からもらう。同じ Sufu遺伝子を持つマウスをホモ接合体、異なる Sufu遺伝子(例えば、野生T396と突然変異したT396Iの異なる Sufu遺伝子)を1つずつもつ個体をヘテロ接合体という。
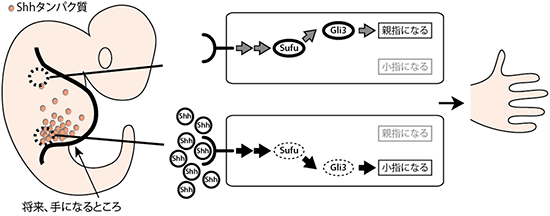
図1 指の向きを決めるメカニズムにおけるHhシグナル伝達系
Hhシグナル伝達系が重要な役割を果たす形態形成の1つに、親指~小指の向きの決定がある。シグナル最上流に位置するソニック・ヘッジホッグ(Shh)タンパク質が将来の小指側からにじみ出し、作られた濃度勾配に従って、小指~親指が作られると考えられている。Sufuタンパク質は、Shhタンパク質からのシグナルを下流に位置するGli転写因子に伝達する。一方で、小指~親指の形成には抑制型のGli3しか関わらず、活性化型のGli1/Gli2は関わらないことが分かっており、これらのよく似た転写因子群(Gli1、Gli2、Gli3)をSufuタンパク質が識別して制御するメカニズムの解明が期待されている。
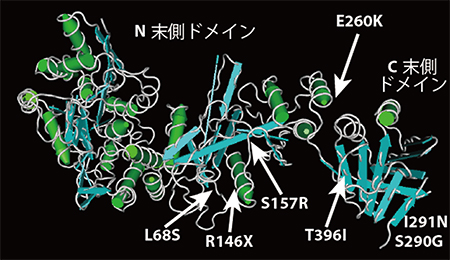
図2 Sufuタンパク質の立体構造とT396Iを含む1アミノ酸置換変異の位置
Sufuタンパク質は、N末側とC末側の2つのドメイン(領域)に別れており、2つのドメインに挟まれた部分に、Gli転写因子が結合すると考えられている注3)。立体構造図の上に、7系統のアミノ酸置換変異の位置を示した。
注3)
Zhang Y. et al. Structural insight into the mutual recognition and regulation between Suppressor of Fused and Gli/Ci. Nat. Commun. 4: 2608 (2013)
Cherry A. et al. Structural basis of SUFU-GLI interaction in human Hedgehog signalling regulation. Acta. Crystallogr. D Biol. Crystallogr. 69: 2563-2579 (2013)
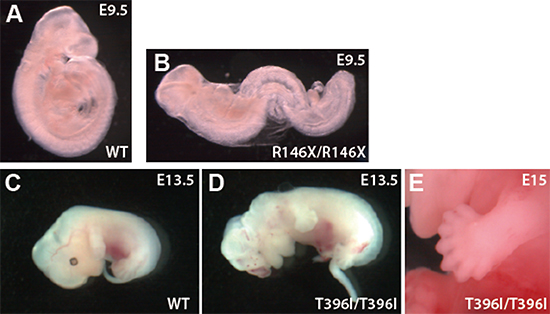
図3 野生型マウス、R146X変異マウス、T396I変異マウスの胚
- A.野生型マウスの発生9.5日の胚
- B.R146X変異マウスの発生9.5日の胚。R146Xは、他の6系統のような1塩基の「置換」とは異なり、1塩基の「欠失」のため、翻訳時に読み枠がずれることにより、機能を完全に失った変異タンパク質を作る。これは、作製方法は一般的なノックアウトとは異なるものの、ノックアウトした場合と同等の現象である。実際に、その結果、R146X変異マウスは、Sufuノックアウトマウスと同等の形態を示し、四肢形態形成が始まる前の発生10日で致死となる。
- C.野生型マウスの発生13.5日の胚
- D.T396I変異マウスの発生13.5日の胚。T396I変異マウスは、顔面や脳神経系の形成不全を起こす。Sufuノックアウトマウスとは異なり、発生後期まで生存するため、T396I変異マウスの解析により、四肢や脳神経の形態形成におけるSufuタンパク質の機能解析が可能になった。
- E.T396I変異マウスの15日胚の左前肢。正常なマウスは5本の指を持っているが、T396I変異マウスは、多指症を起こす。写真の個体は7本の指を持っている。
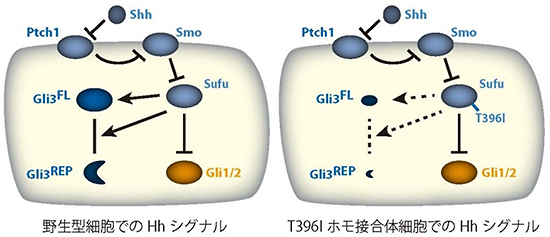
図4 Hhシグナル伝達系のモデルとT396Iタンパク質の活性
Shhタンパク質のシグナルは、Ptch1受容体やSmoタンパク質を介して、Sufuタンパク質に伝達される。これまでの研究から、Sufuタンパク質は、Gli3(Gli3FL)の安定化と、Gli3FLから転写抑制活性を持つGli3REPへの部分的なタンパク質分解反応を促進することが分かっている。また、Sufuタンパク質は、Gli1/Gli2の活性を抑制することも知られている(左)。T396I変異タンパク質は、Gli1/Gli2を制御できる一方、Gli3FL/ REPの制御活性を失っていることが分かった。(右)。すなわち、Gli1/Gli2を抑制する機構とT396を必要とするGli3を制御する機構は、質的に異なった機構であることが明らかになった。