2015年4月28日
理化学研究所
微生物が特定の形の代謝物を作るために必要な酵素遺伝子を発見
-複雑な立体構造の二次代謝物を作るメカニズムの全容解明への第一歩-
要旨
理化学研究所(理研)環境資源科学研究センター天然物生合成研究ユニットの加藤直樹研究員、高橋俊二ユニットリーダーと、同研究センターケミカルバイオロジー研究グループの野川俊彦研究員、長田裕之グループディレクターらの共同研究グループ※は、微生物が特定の立体構造の代謝物を選択的に作るために必要な新しい酵素遺伝子を発見しました。
微生物が作る二次代謝物[1]の中には、ペニシリンやロバスタチン、シクロスポリンなど、有用な生理活性を示すものがあり、私たちはそれらを薬や薬品材料として利用してきました。二次代謝物は、不斉炭素原子[2]を数多く持つ非常に複雑な立体構造をしています。そのような複雑な立体構造の二次代謝物の生合成過程では、二次代謝物の骨格を作る酵素やその骨格を修飾するさまざまな酵素が関わっています。微生物がどのような仕組みで二次代謝物を作り出しているのかを理解することは、二次代謝物の生産性向上や、より有用な二次代謝物を作り出す上で非常に重要です。
共同研究グループは、糸状菌(カビ)の二次代謝物の1つ「エキセチン[3]」に着目しました。そして、その生合成に関わる遺伝子群を同定し、各遺伝子を欠失させた株を作製して遺伝子ごとの機能を解析しました。その結果、ディールス・アルダー(Diels-Alder)反応[4]により作られるエキセチンのデカリン環[5]の立体構造を制御する酵素遺伝子(fsa2)を発見しました。今後、fsa2遺伝子から作られる酵素「Fsa2」がどのようにDiels-Alder反応を制御しているか、そのメカニズムを明らかにすることで、複雑な立体構造を持つ微生物の二次代謝物を選択的に生産するメカニズムの全容解明が進むと期待できます。
本研究は、科学研究費補助金、農林水産業・食品産業科学技術研究推進事業の支援等により実施され、成果は米国の科学雑誌『Biochemical and Biophysical Research Communications』(5月1日号)に先立ち、オンライン版(3月11日付け:日本時間3月12日)に掲載されました。
※共同研究グループ
理化学研究所 環境資源科学研究センター
天然物生合成研究ユニット
研究員 加藤 直樹(かとう なおき)
ユニットリーダー 高橋 俊二(たかはし しゅんじ)
ケミカルバイオロジー研究グループ
研究員 野川 俊彦(のがわ としひこ)
研究嘱託 廣田 洋(ひろた ひろし)
グループディレクター 長田 裕之(おさだ ひろゆき)
韓国生命工学院 ケミカルバイオロジー研究センター
ディレクター Jong Seog Ahn(ジョンソク・アン)
上級研究員 Jae-Hyuk Jang(ゼヒョク・ジャン)
背景
微生物が作る二次代謝物は、不斉炭素原子を多く含む非常に複雑な立体構造をしています。その構造は強力かつ選択的な生物活性と密接な関係があることが知られています。二次代謝物の生合成には、ポリケチド合成酵素(PKS)や非リボソームペプチド合成酵素(NRPS)[6]といった骨格形成に必須な酵素のほか、骨格を修飾するさまざまな酵素が作用することにより、複雑な立体構造になります。こうした多く段階を踏む生合成過程の中で、ある特定の立体構造を選択的に作り出すステップを担う酵素を見つけ、その反応メカニズムを理解することは、医薬品や農薬の素材となりうる有用な二次代謝物を作り出すための第一歩になります。
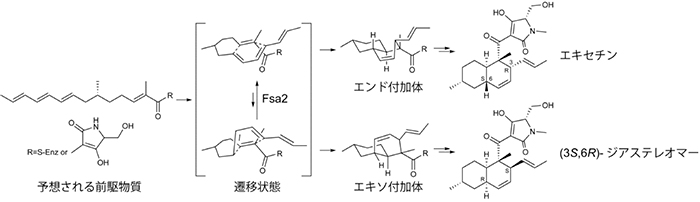
ディールス・アルダー(Diels-Alder)反応は、微生物が作る二次代謝物の生合成において、鍵となる重要なステップです。しかし、その反応を触媒する酵素「Diels-Alderase」は、ごく限られた数の解析例しか報告されていません。そこで、共同研究グループは、ディールス・アルダー反応の関与が指摘されている糸状菌(カビ)の二次代謝物「エキセチン」と「フサリセチンA[3]」(図1)に着目し、その生合成過程の解明に取り組みました。
研究手法と成果
共同研究グループは、エキセチンとフサリセチンAの生合成に関わる遺伝子を同定するため、それら二次代謝物を作る「Fusarium属糸状菌FN080326株」のゲノム配列を解読しました。エキセチンの化学構造から、PKS-NRPSハイブリッド酵素[6]と呼ばれるグループの酵素がエキセチンの骨格形成を担っていることが予想されていました。解読したゲノム配列の中には、PKS-NRPSハイブリッド酵素の遺伝子が1つ(fsa1遺伝子)含まれており、その周辺には、骨格の修飾など、生合成に関わっていそうな遺伝子(fsa2~fsa7遺伝子)が存在していました。そこで、これらの遺伝子をエキセチンの生合成に関わる候補遺伝子として解析を進めました。
まず、二重相同組換え[7]によりfsa1遺伝子を欠失させたfsa1遺伝子欠失株を作製したところ、この株ではエキセチンとフサリセチンAの生産が消失しました。次にfsa1遺伝子欠失株の培養液にエキセチンを添加したところ、フサリセチンAの生産が回復しました。このことから、フサリセチンAはエキセチンを材料に作られる二次代謝物であり、両者の生合成にはfsa1遺伝子が関与していることが分かりました(図1)。
続いて、fsa1遺伝子に隣接するfsa2遺伝子について調べました。fsa2遺伝子から作られるタンパク質のアミノ酸配列は、すでに機能が分かっているタンパク質のいずれのアミノ酸配列とも似ておらず、機能を推定することができませんでした。そこで、fsa2遺伝子欠失株を作製し、その培養液を分析しました。その結果、エキセチンの生産量が大幅に低下することに加え、エキセチンと同じ分子量を示す化合物が新たに検出されました。この化合物を単離、精製し、質量分析および各種二次元核磁気共鳴(NMR)測定を行ったところ、その構造がエキセチンの立体異性体[8]であることが明らかになりました。さらに、旋光度[9]、円偏光二色性(CD)スペクトラム[9]の測定などにより、その立体異性体はデカリン環部分の立体構造に違いがあるジアステレオマー[8]であることを突き止めました(図2)。デカリン環部分は、ディールス・アルダー反応を介して作られることが知られています。このことから、fsa2遺伝子が、ディールス・アルダー反応を介したデカリン環形成における立体構造制御に関わるタンパク質(酵素)を作る遺伝子であることが分かりました。
どのような二次代謝物を作っているのか分からない糸状菌のPKS-NRPSハイブリッド酵素遺伝子の中には、fsa2遺伝子の配列に似た遺伝子がその近傍に存在しているものがあります。今回発見したfsa2遺伝子の機能は、エキセチンに似た二次代謝物の生合成過程において共通したものである可能性が考えられます。
今後の期待
本研究により、微生物が作り出す二次代謝物の生産に関わる新しい酵素遺伝子「fsa2」の存在が明らかとなりました。今後、fsa2遺伝子から作られる酵素「Fsa2」の立体構造を明らかにし、ディールス・アルダー反応のメカニズムの詳細を解明することで、複雑な立体構造の二次代謝物を作り出すメカニズムの全容解明がさらに進むと期待できます。有用な二次代謝物の生合成に関わる微生物の遺伝子の探索と活用は、生合成による物質生産の基盤構築とそれを用いた効率的生産につながる可能性があります。
原論文情報
- Naoki Kato, Toshihiko Nogawa, Hiroshi Hirota, Jae-Hyuk Jang, Shunji Takahashi, Jong Seog Ahn, Hiroyuki Osada, "A new enzyme involved in the control of the stereochemistry in the decalin formation during equisetin biosynthesis", Biochemical and Biophysical Research Communications, doi: 10.1016/j.bbrc.2015.03.011
発表者
理化学研究所
環境資源科学研究センター 天然物生合成研究ユニット
研究員 加藤 直樹(かとう なおき)
ユニットリーダー 高橋 俊二(たかはし しゅんじ)
環境資源科学研究センター ケミカルバイオロジー研究グループ
研究員 野川 俊彦(のがわ としひこ)
研究嘱託 廣田 洋(ひろた ひろし)
グループディレクター 長田 裕之(おさだ ひろゆき)
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.二次代謝物
生物体を構成、維持する上で重要な物質を一次代謝物、生育に必ずしも必須ではない物質を二次代謝物と呼ぶ。微生物において、二次代謝物の生合成に関わる酵素遺伝子はゲノム中のある特定の領域に並んで存在している。 - 2.不斉炭素原子
異なる4つの置換基が共有結合している炭素原子のこと。分子内に複数の不斉原子が含まれている場合、ある分子の鏡像となる立体異性体をエナンチオマー、鏡像ではない立体異性体をジアステレオマーと呼ぶ。二次代謝物の多くは生合成酵素の作用により、ある1つの決まった立体構造が選択的に作り出される。 - 3.エキセチン、フサリセチンA
エキセチン(分子量373)は、フザリウム属糸状菌より単離された二次代謝物。グラム陽性細菌に対する抗菌活性、およびHIV-1インテグラーゼに対する阻害活性を示すことが知られている。一方、フサリセチンA(分子量389)は、フザリウム属糸状菌FN080326株より、がん細胞の形態分化を阻害する物質とした単離された二次代謝物。ユニークな五環性の縮合環構造を有しており、合成化学的にも興味深い化合物である。これまでに複数の有機合成化学者により全合成が報告されている。 - 4.ディールス-アルダー(Diels-Alder)反応、エンド付加体、エキソ付加体
鎖状構造の有機化合物を環状構造に変換する環化反応の中でも、2つ以上の不飽和分子が結合して環状構造をつくる反応(環化付加反応)は、有機化学合成において重要である。ディールス-アルダー反応はその1つで、共役ジエンにアルケン(ジエノファイル)が付加して6員環構造が生じる。生成物はその立体構造に応じて、エンド付加体、エキソ付加体と呼ばれる。多くの二次代謝物の生合成過程において、鍵となるプロセスを担っていることが指摘されているものの、その反応を触媒する酵素「Diels-Alderase」については、ごく限られた数の解析例が報告されているのみである。 - 5.デカリン環
二環性の炭化水素化合物。別名、デカヒドロナフタレン。デカリン環を部分構造に持つ糸状菌由来の二次代謝物としては、高脂血症治療薬であるロバスタチンが有名である。ロバスタチンの骨格構造形成を担うポリケチド合成酵素LovBが、ポリケチド鎖の合成だけでなく、ディールス-アルダー反応にも関わっていることが示されている。類似した酵素によって骨格が作られるエキセチンでも同様のメカニズムによりデカリン環が形成すると推測されてきたため、別の酵素の関与は予想外であった。 - 6.ポリケチド合成酵素(PKS)、非リボソームペプチド合成酵素(NRPS)、PKS-NRPSハイブリッド酵素
微生物の二次代謝物生合成に特徴的な巨大多機能型酵素。ポリケチド合成酵素(PKS)は、アセチルCoAを開始基質に、主にマロニルCoAを伸長基質としてポリケトン鎖を形成する。酵素を構成する触媒ドメインの作用により、還元や脱水、メチル化などの修飾をし、ポリケチド化合物の複雑な骨格を作り出す。非リボソームペプチド合成酵素(NRPS)は、リボソームを介さず、PKSに類似したメカニズムでペプチド鎖を合成する。タンパク質には含まれないアミノ酸を基質とできることも特徴である。また、PKSとNRPSの機能が融合したタイプの酵素も存在し、PKS-NRPSハイブリッド酵素と呼ばれる。糸状菌のPKS-NRPSハイブリッド酵素によって生合成される二次代謝物は、特徴的な化学構造を有しており、さまざまな有用生物活性を示す。 - 7.二重相同組換え
相同組換えとは、染色体上のDNAの塩基配列がよく似た部位(相同配列)同士で入れ替えが起きること。遺伝子組換えは生物が本来持っている機能であり、そのシステムを利用することで人工的に遺伝子を改変することができる。機能を調べたい遺伝子をゲノムから取り除く方法として、その遺伝子の両側で相同組換えを起こさせることで標的の遺伝子を別のDNAと置換する方法が二重相同組換えである。 - 8.立体異性体、ジアステレオマー
同じ分子式で表されるが、構造が異なる物質のことを異性体と呼び、原子の並び順が異なる位置異性体と、原子の並びは同じでも3次元空間での配置の異なる立体異性体に分けられる。右手と左手のように互いに鏡像である立体異性体のことをエナンチオマーと呼び、それ以外の立体異性体をジアステレオマーと呼ぶ。 - 9.旋光度、円偏光二色性(CD)スペクトラム
通常の構造解析で用いられるNMRでは、立体異性体(特にエナンチオマー)の判別が困難であることが多い。そこで、不斉原子に起因する光学活性(旋光性)の違いを利用して異性体の識別がなされる。旋光度やCDの測定はその有効な方法である。
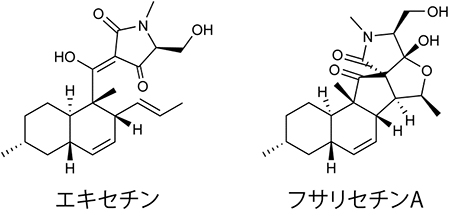
図1 エキセチンとフサリセチンAの化学構造
フサリセチンAは骨格構造がエキセチンと共通していることから、エキセチンを材料として生合成されると予想された。実験の結果、フサリセチンAはエキセチンを材料に作られる二次代謝物であり、両者の生合成にはfsa1遺伝子が関与していることが分かった。