要旨
理化学研究所(理研)多細胞システム形成研究センター染色体分配研究チームの北島智也チームリーダーらの研究チーム※は、卵母細胞[1]の2回の細胞分裂のうち、母方の配偶子(卵子)と父方の配偶子(精子)由来の相同染色体[2]が分配される「減数第一分裂[3]」では、微小管と染色体の正しい接続が不安定になりやすいことを発見しました。
卵母細胞は、減数第一分裂を経て卵子となります。排卵された卵子が受精すると減数第二分裂を完了し、受精卵となって胚発生を開始します。減数第一分裂では、相同染色体が正しく卵子に分配されなくてはなりません。染色体の分配に誤りがあると、流産やダウン症などでみられる染色体数の異常につながります。染色体が正しく分配されるには、微小管が染色体に正しく接続し、その接続が安定する必要があります。微小管は主に染色体上の「動原体[4]」に接続します。体細胞分裂では、微小管が動原体に正しく接続し動原体を引っ張ると、すぐにその接続が安定すると考えられています。しかし、卵母細胞の減数第一分裂では、どのように微小管と動原体の接続が安定するかは、明らかではありませんでした。
研究チームは、マウス卵母細胞の減数第一分裂では、微小管が正しく動原体に接続し動原体を引っ張っても、すぐには微小管と動原体の接続が安定しないことを突き止めました。これは、接続を不安定にする因子であるタンパク質修飾酵素「Aurora B/Cキナーゼ[5]」が動原体近傍に残存することが原因でした。動原体近傍に残存したAurora B/Cキナーゼは、微小管と動原体の正しい接続を不安定にするだけでなく、微小管と動原体の誤った接続の形成に関与していました。
研究チームはさらに、卵母細胞の減数第一分裂では、微小管が動原体を引っ張った後、微小管と動原体の接続が数時間もの長い時間をかけて徐々に安定していくことを発見しました。この安定化は、タンパク質脱修飾酵素の「PP2A-B56フォスファターゼ[5]」が徐々に動原体に集められ、Aurora B/Cキナーゼと拮抗するためだと分かりました。
これらの結果から、卵母細胞の減数第一分裂では、微小管と動原体の正しい接続が不安定となりやすく、体細胞分裂とは異なるメカニズムによって安定することが明らかになりました。また、この特徴的な安定化のメカニズムが、卵母細胞の減数第一分裂で微小管と動原体の接続の誤りが多い原因と関連していることが分かりました。本研究により、卵子をつくるための細胞分裂における染色体分配メカニズムの不安定な一面が見いだされました。
本研究は、米国の科学雑誌『Developmental Cell』(6月8日号)に掲載されるのに先立ち、オンライン版(5月28日付け:日本時間5月29日)に掲載されます。
※研究チーム
理化学研究所 多細胞システム形成研究センター 染色体分配研究チーム
チームリーダー 北島 智也 (きたじま ともや)
研究員 吉田 周平 (よしだ しゅうへい)
元テクニカルスタッフ 海道 雅子 (かいどう まさこ)
背景
細胞は分裂する際に、染色体を娘細胞に正しく分配しなくてはなりません。染色体を分配するために、細胞は微小管を使って染色体を動かします。このとき、微小管は染色体上にある動原体に接続します。微小管が動原体に正しく接続することは、正しい染色体分配の必要条件です。
増殖などのために行われる体細胞分裂では、細胞はDNA複製によりコピーしてできた姉妹染色分体[6]ペアを娘細胞に分配します。その際、姉妹染色分体のそれぞれに動原体が1つずつ形成され、これを姉妹動原体と呼びます。細胞は微小管を姉妹動原体に両方向から接続させ、微小管と動原体の接続を安定させます(図1上)。接続が安定する要因は、姉妹動原体が微小管によって反対方向に引っ張られることにより、接続を不安定にする因子「Aurora Bキナーゼ」と空間的に離れるためと考えられています。このメカニズムにより微小管と動原体が正しく接続し安定することで、体細胞分裂では姉妹染色分体が娘細胞に正しく分配されていると考えられています。
一方、卵母細胞の減数第一分裂では、姉妹染色分体ペアではなく、相同染色体ペアが娘細胞に分配されます。その際、相同染色体のそれぞれに動原体が1つずつ形成され、これを相同動原体と呼びます。減数第一分裂では、微小管は姉妹動原体を一方向から接続し、かつ相同動原体を両方向から接続しなくてはなりません(図1下)。このような特殊なタイプの微小管と動原体の正しい接続が卵母細胞においてどのように安定するのかは、明らかになっていませんでした。
研究手法と成果
研究チームは、マウスを使った実験で卵母細胞の減数第一分裂で微小管と動原体の接続が安定するタイミングを調べました。体細胞分裂では、微小管によって姉妹動原体が正しく引っ張られるのと同時に、微小管と動原体の接続が安定します。ところが卵母細胞の減数第一分裂では、微小管によって相同動原体が正しく引っ張られても、すぐには微小管と動原体の接続が安定しませんでした。そこで研究チームは、「卵母細胞の減数第一分裂では、微小管によって相同動原体が正しく引っ張られても、接続を不安定にする因子が動原体から空間的に離れていないのではないか」と考えました。卵母細胞では、Aurora Bキナーゼに加え、Aurora Cキナーゼが動原体をリン酸化して接続を不安定にすることが知られています(これらを合わせてAurora B/Cと記述)。そこでAurora B/Cの局在を調べたところ、微小管によって相同動原体が正しく引っ張られた後でも、Aurora B/Cは動原体の近傍に残存していました(図2)。
さらに、数時間にわたって微小管と動原体の接続を調べたところ、微小管と動原体の正しい接続はすぐには安定しないものの、長い時間をかけて徐々に安定していくことが分かりました。研究チームは、この安定化は、タンパク質脱修飾酵素の「PP2A-B56フォスファターゼ」が徐々に動原体に集められ、Aurora B/Cと拮抗して動原体を脱リン酸化するためであることを発見しました(図3)。これらの結果から、卵母細胞の減数第一分裂では、微小管と動原体の正しい接続の安定のために体細胞分裂とは異なるメカニズムが働いていることが分かりました。
卵母細胞の減数第一分裂における微小管と動原体の接続は、誤りが多いことが知られています。研究チームは「この誤りが起こる原因は動原体近傍に残存するAurora B/Cではないか」と考えました。この考えを検証するため、微小管によって相同染色体が正しく引っ張られると同時にAurora B/Cの活性を阻害しました。すると、微小管と動原体の正しい接続が安定し、一方で動原体と微小管の誤った接続は起きにくくなりました(図4)。
これらの結果は、卵母細胞の減数第一分裂では、①微小管と動原体の正しい接続がつくられても動原体のリン酸化状態が高く維持されるため、接続が安定しにくくなっている②それが微小管と動原体の誤った接続を起こしやすくしている③体細胞分裂とは異なったメカニズムにより長い時間をかけて動原体を脱リン酸化させ、微小管と動原体の接続を安定させていることを示しています(図5)。
今後の期待
卵母細胞は減数第一分裂を経て卵子となります。排卵された卵子が受精すると減数第二分裂が完了し、受精卵となって胚発生へと進行します。卵母細胞の減数第一分裂の染色体分配は誤りが起きる頻度が高いことが知られています。流産やダウン症などでみられる染色体数の異常の多くがこの誤りに起因しています。今回の研究により、卵母細胞には、染色体分配に重要な微小管と動原体の接続について誤りを起こしやすい性質があることが明らかになりました。正常な卵母細胞では、このような微小管と動原体の接続の誤りは、染色体分配の前までに正しく修正されることが知られていますが、ヒトのような哺乳類の卵母細胞では、染色体分配の誤りの頻度が加齢に伴って上昇することが知られています。このことから卵母細胞は加齢に伴って、動原体と微小管の接続の誤りを修正できなくなっていく可能性が考えられます。さらに研究を進めることで、哺乳類における染色体数の異常の主要な原因が明らかになると期待できます。
原論文情報
- Shuhei Yoshida, Masako Kaido, Tomoya S. Kitajima, "Inherent instability of correct kinetochore-microtubule attachments during meiosis I in oocytes", Developmental Cell
発表者
理化学研究所
多細胞システム形成研究センター 染色体分配研究チーム
チームリーダー 北島 智也 (きたじま ともや)
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.卵母細胞
雌性生殖細胞。ヒトを含む多くの哺乳類では、卵巣の中に蓄えられた卵母細胞が排卵の前に減数第一分裂を完了して卵子となる。排卵された卵子が受精すると減数第二分裂を完了し、受精卵となって胚発生を開始する。 - 2.相同染色体
ほとんどの細胞は、母方の配偶子(卵子)と父方の配偶子(精子)のそれぞれに由来する染色体セットを持っている。例えばヒトでは、23種類の染色体それぞれについて、卵子に由来するものと精子に由来するものの2つがある。これら2つの染色体を相同染色体と呼ぶ。 - 3.減数第一分裂
生殖細胞は配偶子をつくるために、減数第一分裂と減数第二分裂の2回の細胞分裂を連続して行う。体細胞分裂や減数第二分裂では姉妹染色分体(DNA複製によるコピー)が分配されるのに対し、減数第一分裂では相同染色体(母方と父方の配偶子に由来する染色体)が分配される。 - 4.動原体
染色体上に形成されるタンパク質複合体。染色体分配の際に、微小管が染色体を動かすための牽引部位となる。体細胞分裂では姉妹染色分体のそれぞれに動原体が1つずつ形成され、これらを姉妹動原体と呼ぶ。減数第一分裂では相同染色体のそれぞれに動原体が1つずつ形成され、これらを相同動原体と呼ぶ。 - 5.キナーゼ、フォスファターゼ
タンパク質修飾酵素。キナーゼはタンパク質をリン酸化、フォスファターゼは脱リン酸化することでそのタンパク質の活性などを制御する。 - 6.姉妹染色分体
細胞は分裂の前にDNA複製を行う。DNA複製によりできた2つの同一な染色体を姉妹染色分体と呼ぶ。
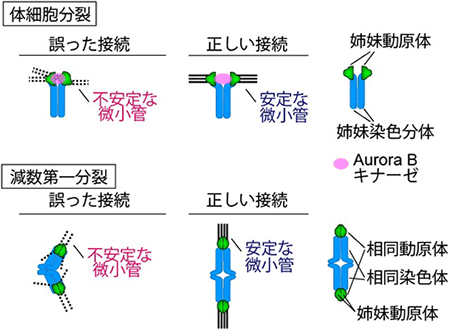
図1 体細胞分裂と減数第一分裂の比較
- 上: 体細胞分裂では、姉妹動原体が正しく微小管と接続し両方向に引っ張られると、動原体がAurora Bキナーゼと空間的に離れ、接続がすぐに安定すると考えられている。
- 下: 卵母細胞の減数第一分裂では、姉妹動原体は一方向から捕らえられ、相同動原体が両方向に引っ張られなくてはならない。このような特殊なタイプの正しい接続が、どのように安定するのかは分かっていなかった。
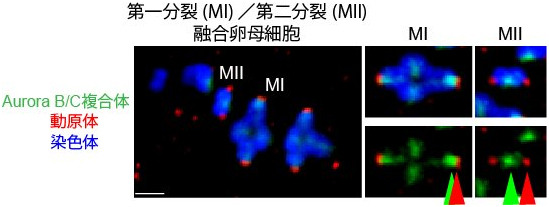
図2 減数第一分裂時と第二分裂時の染色体におけるAurora B/Cの局在を比較
減数第一分裂時のマウス卵母細胞と、減数第二分裂時のマウス卵母細胞を融合させ、Aurora B/C複合体の局在位置を観察した。第一分裂の染色体(MI)では、相同動原体が正しく引っ張られた後でも、動原体(赤)の近傍にAurora B/C(緑)が観察される。これは、第二分裂の染色体(MII)では、動原体からAurora B/Cが遠くはなれているのとは対照的である。
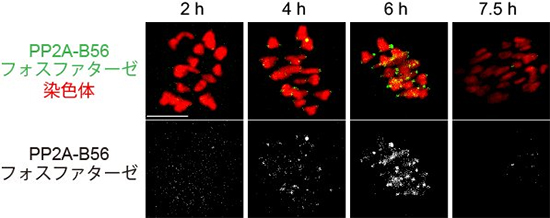
図3 PP2A-B56フォスファターゼが動原体に集まる様子
卵母細胞における微小管と動原体の接続の安定化は数時間かけて徐々に起きる。これは、PP2A-B56フォスファターゼが徐々に動原体に集められ、Aurora B/Cと拮抗するためだった。
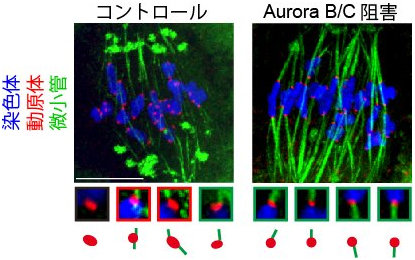
図4 Aurora B/Cの活性を阻害した後の様子
動原体(赤)が微小管(緑)によって正しく引っ張られると同時にAurora B/Cの活性を阻害すると、微小管と動原体の正しい接続が安定し、誤った接続が減少する。
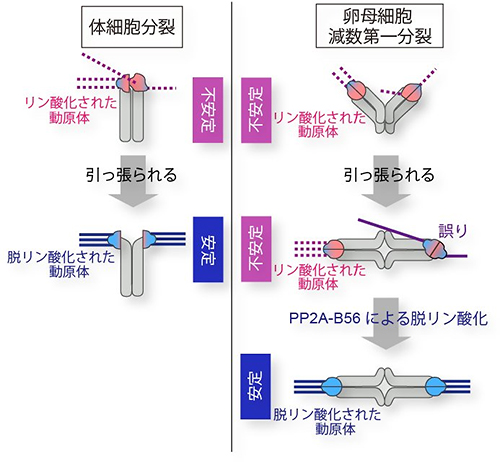
図5 卵母細胞における微小管と動原体の接続の安定化
体細胞分裂では、微小管に動原体が正しく引っ張られるとともに、動原体のリン酸化レベルが減少して微小管と動原体の接続が安定する。一方、卵母細胞の減数第一分裂では、微小管に動原体が正しく引っ張られても、動原体はAurora B/Cにより高いリン酸化レベルが保たれ、微小管と動原体の接続はすぐに安定しない。この際、誤った接続が起きやすくなる。その後、徐々にPP2A-B56フォスファターゼが動原体を脱リン酸化し、動原体と微小管の接続はようやく安定する。