要旨
理化学研究所(理研)多細胞システム形成研究センター形態形成シグナル研究チームの林茂生チームリーダーと大谷哲久テクニカルスタッフらの国際共同研究チーム※は、細胞の伸長を駆動する司令塔の役割を果たす物質が伸長端に正しく配置される仕組みを解明しました。
多細胞生物の体ができる過程では、細胞はそれぞれの機能に応じてさまざまな形に変形します。その中でも細胞の伸長は比較的単純な細胞の形づくりであり、細胞の特定の部位に伸長端が作られ、細胞が一方向に成長することによって起こります。細胞の伸長を駆動するためには、その伸長端に細胞骨格や細胞内輸送などを制御する司令塔のシグナル分子[1]が正しく配置されることが重要です。これまで、形態形成シグナル研究チームはショウジョウバエの剛毛[2]細胞をモデルに細胞伸長の仕組みを調べ、剛毛細胞では「IKKε(アイケイケイイプシロン)」[3]というタンパク質が細胞伸長の司令塔の役割を果たしていることを明らかにしていました。しかし、IKKεが剛毛細胞の伸長端に正しく配置される仕組みは分かっていませんでした。
今回、国際共同研究チームが、その仕組みを調べた結果、IKKεはアダプタータンパク質[4]「Spindle-F(Spn-F)」を介して、細胞質ダイニン[5]と複合体を形成することにより剛毛細胞の伸長端に輸送されていることが分かりました。細胞質ダイニンは、微小管[6]のマイナス端への物質輸送を担う主要な分子モーターです。さらに、伸長端に到達したIKKε複合体は、伸長端で「Javelin-like(Jvl)」[7]と結合することで選択的につなぎとめられていること(係留)を発見しました。以上の結果から、細胞伸長の司令塔は極性輸送[8]と選択的な係留という二段階のプロセスによって伸長端に正しく配置されることが明らかになりました。
本成果は、細胞の多様な形態形成を制御する分子機構の一端を明らかにしただけでなく、細胞内の巧妙な物流システムがいかにして調節・維持されているかを理解する上で重要な知見です。今後、さらに複雑な細胞形態の出来る仕組みや細胞内の物流システムが正しく働く仕組みといった細胞生物学分野における諸問題を解明する手がかりになると期待できます。
本研究は、英国の科学雑誌『Development』のオンライン版(6月19日付け)に掲載されました。
※国際共同研究チーム
理化学研究所 多細胞システム形成研究センター 形態形成シグナル研究チーム
チームリーダー 林 茂生(はやし しげお)
テクニカルスタッフ 大谷 哲久(おおたに てつひさ)
ネゲブ・ベン=グリオン大学 理学部 生命科学科(イスラエル)
准教授 Uri Abdu(ウリ・アブドゥ)
背景
多細胞生物の体ができる過程では、細胞はそれぞれの機能に応じてさまざまな形に変形します。細胞の伸長は、比較的単純な細胞の形づくりであり、神経細胞などさまざまな細胞の形をつくるのに寄与しています。細胞の伸長は、細胞の特定の部位に伸長端が作られ、細胞が一方向に成長することによって起こります。細胞の伸長を駆動するためには、細胞骨格や細胞内輸送などを制御する司令塔のシグナル分子が、伸び続ける細胞の伸長端に常に正確に配置されることが重要です(図1)。しかし、これまで細胞伸長の司令塔がどのように伸長端に配置されるかは良く分かっていませんでした。
これまで形態形成シグナル研究チームは、機械刺激や化学刺激を受容する外感覚器として働く、ショウジョウバエの剛毛細胞に注目して細胞伸長の仕組みを調べてきました。毛のように細長い形の剛毛細胞は、さなぎ期には1日で350マイクロメートル(μm、1μmは100万分の1メートル)も伸長することが知られています。研究チームは、この細胞の形成過程では「IKKε(アイケイケイイプシロン)」というタンパク質が細胞の伸長端に局在し、伸長の司令塔の役割を果たしていることを明らかにし、2011年2月に発表しました。しかし、IKKεが伸長端に正しく配置される仕組みについては分かっていませんでした。
研究手法と成果
国際共同研究チームは、IKKεと複合体を形成することが知られているタンパク質「Spindle-F(Spn-F)」に着目しました。spn-F遺伝子が変異したショウジョウバエ(spn-F遺伝子変異体)の剛毛細胞を調べたところ、伸長端にIKKεが配置されず(図2)通常より短く枝分かれした剛毛となり、剛毛の形づくりが正しくできませんでした(図3)。
さらに、Spn-Fが他のタンパク質と結合するかを生化学的に調べたところ、細胞質ダイニンと複合体を形成することが分かりました。細胞質ダイニンは、微小管のマイナス端に物質を運ぶのに重要な働きをする分子モーターです。このことから、Spn-FはIKKεと細胞質ダイニンの結合を介在するアダプターとして働いていることが明らかとなりました(図4)。
次に、伸長端においてSpn-Fがどの程度安定的に存在するかを調べるため、Spn-Fを緑色蛍光タンパク質(GFP)で標識し、光褪色後蛍光回復法(FRAP法)[9]によって、伸長端のSpn-Fが細胞質のSpn-Fと置き換わる速度を調べました。FRAP法は、レーザー照射などにより細胞内の特定の領域の蛍光を褪色させた後にその領域における蛍光の回復を観察することにより、蛍光分子の動態に関する情報を取得できる手法です。観察の結果、Spn-Fはいったん伸長端に配置されると置き換わりはおきず、安定に伸長端につなぎとめられていることが分かりました(図5)。
続いて、研究チームはIKKε-Spn-F複合体が伸長端につなぎとめられる仕組みを調べました。その結果、Spn-Fと結合することが知られているJavelin-like(Jvl)というタンパク質がIKKε-Spn-F複合体をつなぎとめるために重要であることが分かりました。jvl遺伝子が変異したショウジョウバエ(jvl遺伝子変異体)の剛毛細胞では、IKKε-Spn-F複合体は細胞伸長の初期では伸長端に存在しましたが、伸長が進むにつれて伸長端から失われていました(図6)。
さらに、蛍光タンパク質で標識したSpn-FとJvlの動きを培養細胞においてライブ・イメージングによって調べたところ、Spn-FとJvlがそれぞれ単独で存在する時には微小管に沿ってダイナミックに動きながら輸送されました。これに対し、Spn-F-Jvl複合体が形成されると動かないことが分かりました。JvlとIKKε-Spn-F複合体はそれぞれ微小管に沿って伸長端へと輸送され、先端部に蓄積・濃縮されることで複合体を形成しやすくなると考えられます。このことから、Spn-FとJvlが結合することによってその動きを止め、伸長端につなぎとめることによって、IKKεが安定的に伸長端に配置されていると考えられました。
これらの結果から、ショウジョウバエの剛毛細胞では、細胞伸長の司令塔であるIKKε-Spn-F複合体が、極性輸送と選択的な係留という2段階のステップによって伸長端に配置されていることが明らかになりました(図7)。
今後の期待
今回、ショウジョウバエの剛毛細胞における細胞伸長の司令塔であるIKKεが極性輸送と選択的な係留という2段階のプロセスによって伸長端に配置されていることが明らかになりました。この結果は、さらに複雑な形態をとる細胞形成がどのような仕組みで制御されているかを理解するうえで重要な成果です。
また、形態形成シグナル研究チームはこれまで細胞質ダイニンによって伸長端に運ばれる別の分子が、細胞伸長の伸長端に留まるIKKεとは異なり、伸長端と細胞体の間を往復運動していることを見いだしています。これらの結果は剛毛細胞の伸長端が細胞質ダイニンの積荷の仕分けを行う物流拠点として働いていること、すなわち伸長端において積荷を細胞体に送り返すのか、それとも係留するのかという選択がなされていることを示しています。今回の成果は、細胞内でどのように物流拠点が形成され維持されるのか、またどのように積荷の仕分けが行われるのかといった細胞生物学分野における諸問題を解明する手がかりになると期待できます。
原論文情報
- Tetsuhisa Otani, Kenzi Oshima, Akiyo Kimpara, Michiko Takeda, Uri Abdu, Shigeo Hayashi, "A transport and retention mechanism for the sustained distal localization of Spn-F-IKKε during Drosophila bristle elongation", Development, doi: 10.1242/dev.121863
発表者
理化学研究所
多細胞システム形成研究センター 形態形成シグナル研究チーム
チームリーダー 林 茂生(はやし しげお)
テクニカルスタッフ 大谷 哲久(おおたに てつひさ)
 林 茂生
林 茂生
 大谷 哲久
大谷 哲久
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.シグナル分子
細胞の中の情報伝達に関わる分子。タンパク質リン酸化酵素やGTP結合タンパク質などが知られる。細胞内の情報伝達系においては、シグナル分子間で酵素反応が次々に起こることによって情報が受け渡され、細胞の運命や振る舞いなどが決定する。 - 2.剛毛
ショウジョウバエをはじめとする昆虫の体表に存在する毛のような構造で、機械刺激や化学刺激を受容する感覚器として働く。1つの細胞が伸長した後に、キチン質を主成分とするクチクラを分泌することによって形成される。 - 3.IKKε(アイケイケイイプシロン)
進化的に保存されたタンパク質リン酸化酵素であり、脊椎動物においてはがんや自然免疫、あるいは神経変性疾患などに関与すると考えられている。ショウジョウバエでは剛毛細胞の形づくりのほか、体軸形成においても重要な働きをすることが知られている。 - 4.アダプタータンパク質
タンパク質同士の結合を介在するタンパク質。通常は互いに強く結合することができないタンパク質同士の結合を介在することにより、細胞内の情報伝達を助けたりタンパク質の複合体形成を助けたりする。Spindle-Fはアダプタータンパク質の1種であり、ショウジョウバエの体軸形成において働く因子として同定された。IKKεなどいくつかのタンパク質と結合することが知られているが、これまで詳細な分子機能は不明であった。 - 5.細胞質ダイニン
微小管のマイナス端に向かって動く分子モーター。細胞内でさまざまな積荷を微小管のマイナス端に向かって輸送するために使われる。 - 6.微小管
細胞の中に存在する細胞骨格の1種であり、チューブリンタンパク質の重合によって形成される繊維状の構造。細胞内輸送において分子モーターの足場となる。微小管繊維の両端は性質が異なり、チューブリン2量体が付加されやすい末端をプラス端、解離しやすい末端をマイナス端と呼ぶ。 - 7.Javelin-like
ショウジョウバエの剛毛形成に関わる遺伝子として同定されたアダプタータンパク質。 javelin-like変異体においては剛毛の形状が直線的になり、先端部が肥大して槍(Javelin)のように見えることから命名された。微小管と結合することが知られているが、詳細な分子機能は不明であった。 - 8.極性輸送
細胞の特定の部位に向かって方向性をもって物質が輸送されること。細胞の非対称性を作り維持するのに重要な役割を果たす。 - 9.光褪色後蛍光回復法(FRAP法)
FRAPはFluorescence Recovery After Photobleachingの略。蛍光分子に強い励起光を与えると不可逆的に蛍光が褪色することを利用して、細胞内の特定の領域における蛍光分子の動態を調べる方法。細胞内の特定の領域の蛍光を褪色させた後にその領域における蛍光の回復を観察することにより、蛍光分子の動態に関する情報を取得することができる。

図1 細胞伸長の司令塔
細胞が伸長する時には、細胞の伸長端にシグナル分子が集積し、司令塔の役割を果たして細胞骨格や細胞内輸送を協調的に制御することによって、一方向への細胞の成長が促進される。
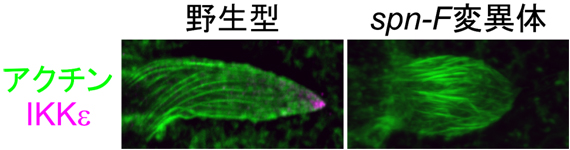
図2 野生型とspn-F遺伝子変異体の剛毛細胞におけるIKKεの局在
- 左: 野生型。IKKε(ピンク)が伸長端に集積している。
- 右: spn-F遺伝子変異体。IKKεの伸長端への集積が消失している。
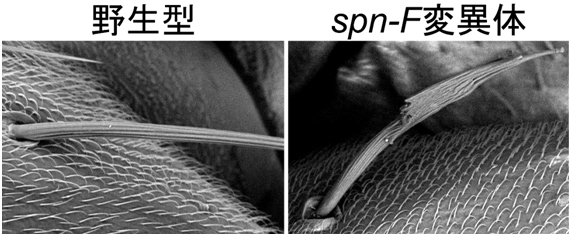
図3 spn-F遺伝子変異体で認められる剛毛の形態異常
spn-F遺伝子変異体の剛毛は直線的ではなく、短く枝分かれした形状となっていた(右)。これは、IKKεをうまく伸長端に配置できないため、細胞骨格や細胞内輸送に異常を来たし、細胞の秩序だった成長が破綻したためと考えられる。
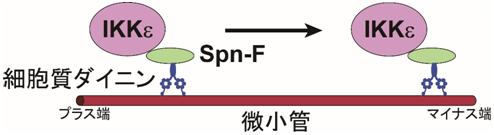
図4 Spn-FがIKKεと細胞質ダイニンをつなぐアダプターとして働く様子
Spn-FはIKKεおよび細胞質ダイニン(微小管マイナス端への輸送を担う分子モーター)と複合体を形成する。このことから、Spn-FはIKKεと細胞質ダイニンをつなぐアダプターとして働いていることが分かった。

図5 Spn-Fタンパク質が伸長端に係留される様子
- 左: 伸長端(ピンク枠)にレーザーを照射した画像。伸長端のSpn-Fの蛍光はレーザー照射による褪色後、回復しない。このことから、伸長端のSpn-Fが蛍光を褪色していない細胞質のSpn-Fと置き換わっていないことが分かる。(細胞質のSpn-Fタンパク質は全体に分布し、密度が低いため蛍光が分かりづらい)
- 右: 細胞質だけ(ピンク枠)にレーザーを照射した画像。蛍光を褪色した細胞質のSpn-Fが、蛍光を褪色していない伸長端のSpn-Fと置き換わった場合、伸長端部分は暗くなるはずだが、伸長端のSpn-Fタンパク質は蛍光を発し続けている。(細胞質のSpn-Fタンパク質は全体に分布し、密度が低いためレーザー照射による褪色が分かりづらい)
これらの観察結果からSpn-Fタンパク質は伸長端に安定的につなぎとめられていることがわかった。
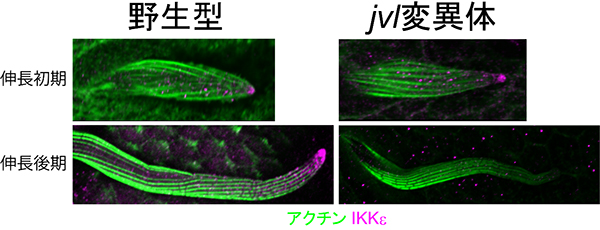
図6 jvl遺伝子変異体で確認されたIKKeの係留の異常
- 上: jvl遺伝子変異体の剛毛細胞では、細胞伸長の初期においてはIKKεが正常に伸長端に配置されていた。
- 下: jvl遺伝子変異体の剛毛細胞では細胞伸長が進行するにつれ、IKKεは徐々に伸長端から失われ、伸長後期には伸長端への集積が消失した。
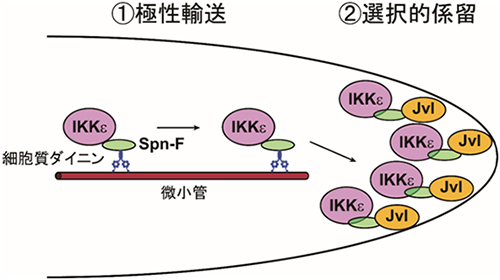
図7 IKKεが伸長端に正しく配置される仕組み
剛毛細胞の伸長の司令塔であるIKKεは、2段階のプロセスによって伸長端に正しく配置される。①まず、IKKεはアダプタータンパク質Spn-Fを介して微小管マイナス端への輸送を担う分子モーターである細胞質ダイニンと複合体を形成し、伸長端へと輸送される(極性輸送)。②伸長端に到達したIKKε-Spn-F複合体はJvlの働きによって伸長端につなぎとめる(選択的係留)。