要旨
理化学研究所(理研)バイオリソースセンター遺伝工学基盤技術室の小倉淳郎室長、畑中勇輝特別研究員らの共同研究チーム※は、マウスにおいて着床前に発現すると死に至るレトロトランスポゾンについて、発現を抑制・制御する仕組みを明らかにしました。
ゲノム上のレトロトランスポゾンは活性化し転写されることでRNAとなり、さらに逆転写酵素[1]によりDNAに戻ることでゲノム中を移動し、コピー数を増やしていきます。哺乳類のゲノムにはレトロトランスポゾンのような反復配列が多く含まれており、全体の約40%を占めています。このことから発生過程において、これらレトロトランスポゾンの抑制はゲノム情報を保護するためにも必須であり、ゲノムワイドな制御が必要です。通常、レトロトランスポゾンの抑制はDNAメチル化[2]によって制御されていますが、着床前初期胚はゲノムワイドにDNAの低メチル化状態であるため、その他の制御が考えられていました。しかし、実験的に確認した例はありませんでした。
共同研究チームは、着床前初期胚においてゲノムの足場であるヒストン[3]がダイナミックに置換される現象に着目し、これを制御するタンパク質「CAF-1」の機能とレトロトランスポゾン制御の関係を調べました。
その結果、CAF-1をノックダウン[4]したマウスの着床前初期胚(CAF-1KD胚)において、レトロトランスポゾンの抑制が効かなくなり(脱抑制)胚性致死となることが分かりました。このCAF-1KD胚に逆転写酵素阻害剤を添加すると胚の生存率が大幅に改善することから、CAF-1をノックダウンさせたことによるヒストン置換の阻害がレトロトランスポゾンの脱抑制を導き、これが胚性致死を引き起こすことを見いだしました。さらに解析を進めていくと、CAF-1による抑制性ヒストン修飾[5]であるH3K9me3とH4K20me3の蓄積がレトロトランスポゾンの抑制に重要であることを突き止めました。
これら研究成果によりレトロトランスポゾンからゲノムの品質管理機構としてDNAメチル化だけでなくヒストン置換を介した機構の存在を明らかにしました。この成果は、哺乳類特有に進化した遺伝子発現プログラムの理解に繋がることが期待されます。
本研究は、11月2日の週に米国アカデミー紀要『Proceedings of the National Academy of Sciences of the United States of America(PNAS)』に掲載されます。
※共同研究チーム
理化学研究所 バイオリソースセンター 遺伝工学基盤技術室
室長 小倉 淳郎(おぐら あつお)
特別研究員 畑中 勇輝(はたなか ゆうき)
九州大学 稲盛フロンティア研究センター
教授 束田 裕一(つかだ ゆういち)
九州大学大学院 医学研究院
准教授 大川 恭行(おおかわ やすゆき)
東北大学大学院 医学系研究科
講師 児玉 栄一(こだま えいいち)
背景
遺伝子情報であるDNAがヒストンに巻かれた構造をヌクレオソームといい、その基本単位がつながったものをクロマチン構造といいます。遺伝子発現は、ヒストンの化学的修飾 (エピジェネティック修飾[6])やヒストンの置換によりクロマチン構造を変化させることで調節されています。
レトロトランスポゾンはゲノム上に存在しており、活性化・転写されることでRNAとなり、さらに逆転写酵素によりDNAに戻ることでゲノム中を移動し、コピー数を増やしていきます。ゲノム情報を保護するためにも、これらレトロトランスポゾンの抑制は必須であり、ゲノムワイドな転写制御が必要です。
転写制御に密接に関連するヒストンH3にはバリアント[7]が存在し、一般的に転写制御においてH3.1/3.2は抑制性、H3.3は転写活性に関係しています。このヒストンバリアントを置換する役割を持つタンパク質はヒストンシャペロン[8]と呼ばれ、これまでにさまざまなヒストンシャペロンが同定されています。この中で「CAF-1」と呼ばれるヒストンシャペロンは、抑制性のヒストンバリアントH3.1/3.2のヌクレオソームへの取り込みに必要とされています。これまでの研究により、CAF-1が欠損すると、着床前に胚性致死となることが報告されていますが、共同研究チームは、これはCAF-1の欠損が、H3.1/3.2の欠損を引き起こし、レトロトランスポゾンの脱抑制につながっているからではないかと考えました。
研究手法と成果
共同研究チームは、塩基配列を変えることなく遺伝子発現を抑制するRNA干渉法によるノックダウン法で、マウスのCAF-1ノックダウン胚(CAF-1KD胚)を作製し、はじめに胚発生を観察しました。その結果、CAF-1KD胚は、これまでの研究で言われていた通り桑実期までは発生しますが、着床前である胚盤胞期までに致死となることが観察されました(図1)。
次に桑実期胚におけるレトロトランスポゾン (LINE-1、SINE-B2、及びIAP)の発現を調べたところ、通常の胚と比べCAF-1KD胚では顕著に増加していることが分かりました。
このレトロトランスポゾンの脱抑制と胚性致死との関係を明らかにするために、レトロトランスポゾンの逆転写を阻害する逆転写酵素阻害剤の添加が胚発生に及ぼす影響を調べました。その結果、逆転写酵素阻害剤によりCAF-1KD胚の胚盤胞期への生存率が大幅に改善することが分かりました。また、CAF-1KD胚においてレトロトランスポゾン領域のヒストンバリアントの蓄積量を調べると、予想通り抑制性のH3.1/3.2が少なく転写活性のH3.3が豊富であり、さらにH3.1に豊富な抑制性ヒストン修飾のH3K9me2、H3K9me3、H3K27me3、及びH4K20me3が減少していることが明らかになりました。
この抑制性ヒストン修飾の減少とレトロトランスポゾンの脱抑制の関係を明らかにするために、これら修飾酵素をノックダウンした胚を作製し、胚発生の観察と桑実期胚におけるレトロトランスポゾンの発現を調べました。その結果、H3K9me3を修飾する酵素であるESET及びH4K20me3酵素であるSuv420h1/2をノックダウンした胚において胚生存率が顕著に減少し、レトロトランスポゾンの脱抑制が認められました。これはH3K9とH4K20をメチル化させない変異体であるH3K9RとH4K20Rを作用させることで、H3K9とH4K20の機能を阻害するドミナントネガティブ実験[9]でも同様の結果が得られました。これらのことから、抑制性ヒストン修飾であるH3K9me3とH4K20me3がレトロトランスポゾンの抑制に重要であることを突き止めました。
本研究結果により、ヒストンシャペロンCAF-1によるヒストン置換により抑制性ヒストン修飾が蓄積され、これが着床前初期胚のDNA低メチル化状態におけるレトロトランスポゾンの発現制御に必須であることが明らかになりました(図2)。
今後の期待
精子及び卵子は受精後エピジェネティックなリプログラミングを経ることで、分化全能性を獲得した受精卵となります。このリプログラミングでは、遺伝子発現の抑制に必須なDNAメチル化がゲノムワイドに外れることが知られています。しかし、レトロトランスポゾンや体細胞など各細胞の形成に必須な遺伝子群は抑制されなければなりません。本研究結果により、哺乳類初期胚においてヒストンシャペロンによるヒストン置換を介したダイナミックな発現抑制機構が初めて明らかになりました。
今後、このリプログラミング過程においてヒストンシャペロンとその下流制御遺伝子群との関係性を明らかにできれば、哺乳類特有である着床前胚の分化制御機構の解明だけでなく、胚性幹(ES) 細胞などが持つ分化多能性獲得や体細胞への分化制御機構の解明に繋がることが期待できます。
原論文情報
- Yuki Hatanaka, Kimiko Inoue, Mami Oikawa, Satoshi Kamimura, Narumi Ogonuki, Eiichi N. Kodama, Yasuyuki Ohkawa, Yu-ichi Tsukada, Atsuo Ogura, "Histone chaperone CAF-1 mediates repressive histone modifications to protect preimplantation mouse embryos from endogenous retrotransposons", Proceedings of the National Academy of Sciences U.S.A., doi: 10.1073/pnas.1512775112
発表者
理化学研究所
バイオリソースセンター 遺伝工学基盤技術室
室長 小倉 淳郎(おぐら あつお)
特別研究員 畑中 勇輝(はたなか ゆうき)
 畑中 勇輝
畑中 勇輝
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.逆転写酵素
RNA依存性DNAポリメラーゼ。DNAの転写産物であるRNAを読み取ってDNAに転写する(逆転写)する酵素。 - 2.DNAメチル化
DNAを構成するシトシン(C)のメチル化。連続したシトシンとグアニン(G)の配列CGのシトシンがメチル化される。DNAメチル化された遺伝子は転写抑制となる。 - 3.ヒストン
DNAが存在する核において、DNAを格納しているタンパク質のこと。ヒストンはH2A、H2B、H3及びH4がそれぞれ二分子ずつ集合し8量体を形成し、これにDNAが巻き付いている(ヌクレオソーム)。 - 4.ノックダウン
遺伝子を破壊するノックアウトと異なり、特定の遺伝子を破壊せずにその転写産物を減少させることで、その遺伝子の機能を阻害する方法。 - 5.抑制性ヒストン修飾
遺伝子の転写に抑制的に働くヒストンのメチル化修飾。ヒストンH3やH4のN末端領域で修飾され、ヒストンH3のN末端から9番目リジン残基のジ/トリメチル化 (H3K9me2/3)、H3K27me3、さらにH4K20me3などが抑制性ヒストン修飾として知られる。これら修飾は凝集されたクロマチン構造を形成するヘテロクロマチン (転写抑制性)に豊富に蓄積される。 - 6.エピジェネティック修飾
DNAやヒストンに修飾される化学的修飾。主にDNAはメチル化、ヒストンはメチル化やアセチル化が付加され、これが遺伝子発現の調節に働いている。これら修飾は様々な酵素により修飾され、特異的な修飾パターンにより細胞特異的な遺伝子発現プロファイルが構築される。 - 7.バリアント
タンパク質のアミノ酸配列は類似しているが、数アミノ酸程度異なっている変異体のこと。タンパク質の機能はほとんど変わらないが、ヒストンバリアントの場合には、バリアントにより遺伝子発現制御が行なわれる。 - 8.シャペロン
ヒストンに結合し、ヒストンがヌクレオソーム(DNAとヒストン)に会合もしくは脱離するのを補助するタンパク質のこと。 - 9.ドミナントネガティブ
目的遺伝子の正常なタンパク質に対し変異タンパク質をドミナント(優位)に作用させることで,目的タンパク質の機能を阻害すること。
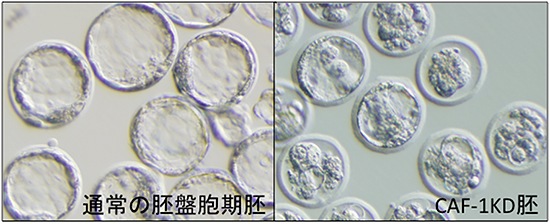
図1 CAF-1KD胚における胚性致死
胚盤胞期となる受精後4日目の写真。左が通常の胚、右はCAF-1KD胚。
CAF-1KD胚は胚盤胞期(受精後4日)までに致死となることが観察された。
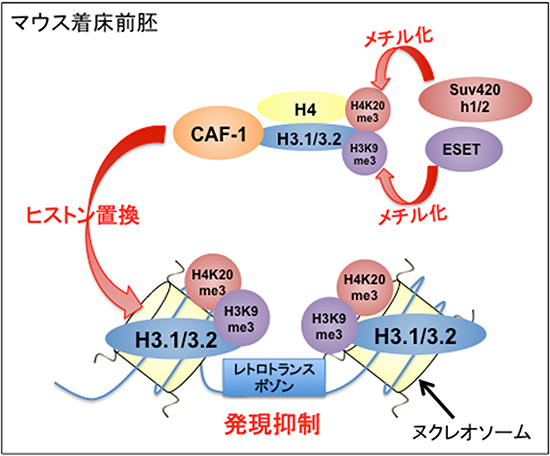
図2 CAF-1によるレトロトランスポゾン抑制機構の模式図
ヒストンシャペロンCAF-1によるヒストン置換により抑制性ヒストン修飾が蓄積され、これが着床前初期胚のDNA低メチル化状態におけるレトロトランスポゾンの発現制御に必須であることが明らかになった。