要旨
理化学研究所(理研)生命システム研究センター発生動態研究チームの大浪修一チームリーダーと髙山順研究員の研究チームは、線虫C. elegans[1]の受精の際に精子のカルシウム透過性チャネル[2]が卵子の中に「受精カルシウム波[3]」を引き起こすことを明らかにし、精子が卵子を活性化する新しい仕組みを解明しました。
動物の一生は、精子と卵子が受精することから始まります。卵子は物質の合成をほとんど行わない不活発な細胞ですが、精子と受精すると活発に物質を合成し、細胞分裂を始める胚へと状態が大きく転換します。これを「卵子の活性化」と呼びます。この転換のきっかけとなるのが、卵子内のカルシウム濃度変化が伝播していく現象「受精カルシウム波」です。
研究チームは、精子が受精カルシウム波をどのように引き起こしているかを明らかにするため、体が透明かつ遺伝学的実験が容易な線虫C. elegansを用いて、その受精カルシウム波を高速イメージングと画像処理によって捉えました。さらに遺伝学的実験とシミュレーションを組み合わせて解析を行い、精子に存在する「TRP-3」というカルシウム透過性チャネルが、受精直後に精子侵入点付近で急激なカルシウム濃度の上昇を引き起こし、これがきっかけとなって卵子全体に伝播するカルシウム波が発生することを発見しました。受精カルシウム波を引き起こす仕組みは、生物種によって異なると考えられています。今回発見した仕組みは、精子のカルシウム透過性チャネルが細胞外から卵子にカルシウムイオンを流入させる「精子導管仮説」を支持するものです。
本研究によって、受精カルシウム波を定量的かつ遺伝学的に解析できる実験系が確立されました。今後、この実験系を活用することで、受精による卵子の状態転換の仕組みの包括的解明につながると期待できます。
本研究の一部は、日本学術振興会科学研究費補助金の助成によって行われました。また、動画像データの定量化は、科学技術振興機構ライフサイエンスデータベース統合推進事業(統合化推進プログラム)の一環として実施されました。さらには、本成果に関するデータの記述手法や、今後データを掲載するデータベースSSBDは同プログラムの一環として開発されたものです。
成果は、米国の科学雑誌『Cell Reports』(4月19日号)に掲載されるのに先立ち、オンライン版(4月7日付け:日本時間4月8日)に掲載されます。
背景
動物の一生は、卵子と精子が受精することから始まります。卵子は物質の合成をほとんど行わない不活発な細胞ですが、受精を機に、それまで抑制されていた物質の合成や細胞分裂の準備を開始し、活発で全能性[4]を持った細胞へと状態が大きく転換します。これを「卵子の活性化」と呼び、自然にみられる全能性獲得への一過程として注目されています。
卵子の状態が大きく転換するきっかけとなるのが、「受精カルシウム波」です。受精カルシウム波は、卵子内のカルシウム貯蔵庫[5]である小胞体などから細胞質へ連鎖的にカルシウムイオン(Ca2+、以下イオンは省略)が放出されることで生じます。受精カルシウム波は、受精という卵子のごく一部分(精子侵入点)で生じた出来事を卵子全体に伝え、その後の卵子の活性化反応を開始するよう指令すると考えられています。精子がどのようにして受精カルシウム波を引き起こすかについては、主に3つの仕組みが提唱されており、生物種によって異なる仕組みが使われていると考えられています。複数の仕組みが使われている可能性も考えられます。
第1の仕組みは、精子内に含まれる酵素などの可溶性因子が、受精により卵子へ受け渡され、卵子内で一連の化学反応が起こり、その結果、卵子内の小胞体からのカルシウム放出を誘導するというものです。可溶性因子として、タンパク質「PLC-zeta」が同定されています。この仕組みは、ほ乳類を中心に広く研究されています。
第2の仕組みは、精子の細胞表面にあるリガンド(特定の受容体に特異的に結合する物質)と、卵子の細胞表面にある受容体が相互作用することで、卵子内に化学反応が起こり、卵子内の小胞体からのカルシウム放出を誘導するというものです。この仕組みは、アフリカツメガエルを中心に研究が進んでいます。
第3の仕組みは、「精子導管仮説」です。精子の細胞膜にカルシウム透過性チャネルが存在すると仮定します。受精により精子の細胞膜と卵子の細胞膜が融合し、精子のカルシウム透過性チャネルが導管(通り道)として働くことで、卵子内に細胞外からカルシウムを流入させます。この流入したカルシウムが、卵子内のカルシウム貯蔵庫からのさらなるカルシウム放出を誘導するというものです。精子導管仮説は、受精カルシウム波の発見者の1人であるライオネル・ジャッフェ博士が1991年に提唱したものです。しかしこれまで、実験的根拠に乏しく、どの生物種においても精子の細胞膜上で導管として働くカルシウム透過性チャネルが見つかっていないことから有力な仮説とは見なされてきませんでした。
研究手法と成果
これまで受精カルシウム波の研究の多くは生化学的に解析され、遺伝学的な解析はあまり行われてきませんでした。そこで、研究チームはモデル生物として、遺伝学的操作が容易で雌雄同体の線虫C. elegansを用いました(図1A)。C. elegansは、体が透明で生きたまま細胞内を顕微鏡観察することができますが、受精は非常に素早い過程であるため、詳細な観察がほとんど行われていませんでした。
研究チームは、蛍光カルシウム指示薬[6]と高速イメージングが可能なスピニングディスク共焦点顕微鏡[7]を用いて、C. elegansの受精直後の卵子のカルシウム濃度変化を動画撮影しました。C. elegansの受精は体内で排卵直後に起こり、視野内を卵子が移動するため、卵子内のカルシウム濃度変化の空間的な特徴がうまく捉えられません。そのため、画像処理技術を用いて卵子領域のみを切り分け、一定の形に整形しました(図1B)。その結果、受精直後に精子侵入点付近で急激にカルシウム濃度が上昇し、その後卵子全体に伝播するという二相性の波形を示すことが分かりました。研究チームは2つの波形を「局所波」および「大域波」と名付けました。
精子侵入点付近で急激にカルシウム濃度が上昇することから、研究チームは精子のカルシウム透過性チャネルが関係しているのではないかと考えました。C. elegansの精子には「TRP-3」というカルシウム透過性チャネルが存在することが知られています。そこで、精子のTRP-3チャネルを欠損させた変異体(trp-3変異体)を用いて受精を観察しました。その結果、trp-3変異体では局所波は生じないことが分かりました(図2)。一方で、大域波は遅れて生じることも分かりました。したがって、TRP-3チャネルは局所波の発生のみならず、大域波が発生するタイミングも決めていることが明らかになりました。
これらの結果は、精子のTRP-3チャネルが受精カルシウム波、特に局所波の発生に関わることを示しています。しかし、本当にTRP-3チャネルが細胞外からカルシウムを流入させているのか、この実験結果だけでは分かりません。
精子のカルシウム透過性チャネルを利用する別の仕組みとしては、受精前に精子内にカルシウムを溜め込み、融合とともに卵子内に放出するという「カルシウム爆弾仮説」も考えられます(図3A)。そこで研究チームは、受精直後の精子内のカルシウム濃度変化を観測し、カルシウム爆弾仮説に基づくカルシウム濃度変化のシミュレーション結果と比較しました(図3B)。観測の結果、受精直後の精子内のカルシウム濃度は卵子と同程度であり、受精後に上昇していました。カルシウム爆弾仮説では高濃度からの急激な低下が予測されるため、観測結果とは一致しません。このことからカルシウム爆弾仮説よりも、精子導管仮説の方が有力であることが示されました。
もし精子導管仮説が正しいとすると、C. elegansの受精は精子と卵子の直接の膜融合という方式で行われ、細胞外からカルシウムを取り込むためにTRP-3チャネルを含む精子の細胞膜が、細胞外領域と面した状態が生じなければなりません。そこで研究チームは、精子のTRP-3チャネルに赤色の蛍光タンパク質「TagRFP-T」を融合させ蛍光標識しました(図4A)。同時に卵子の細胞膜を「GFP::PH」という蛍光タンパク質で緑色に蛍光標識した株を用いて、これらの受精をスピニングディスク共焦点顕微鏡で観察しました(図4B)。その結果、C. elegansの受精は確かに精子と卵子の直接の膜融合で行われること、またその際に精子のTRP-3チャネルは細胞外領域に面した状態でいることが分かりました。
以上の結果は、どれも精子導管仮説が予測することと一致します。したがって、C. elegansの受精時に見られる局所波は精子のTRP-3チャネルが導管として働くことで引き起こされていると考えられます。
今後の期待
本研究により、受精カルシウム波を引き起こす新しい仕組みが発見されました。これは受精のみならず、細胞同士が融合する際に、その中身ではなく細胞膜成分を受け渡すことで、情報を伝えるという方式があることを示唆しています。細胞自体の融合や分泌小胞の融合における新たな細胞間情報伝達の仕組みが、今後明らかになるかもしれません。
また本研究を通じて、C. elegansにおける受精および受精カルシウム波の高速イメージング手法と受精カルシウム波の定量的な解析手法が確立されました。C. elegansはさまざまな遺伝学的実験が容易に適用できるため、これらを組み合わせることで受精および卵子の活性化の仕組みがより深く理解されると期待できます。
原論文情報
- Jun Takayama and Shuichi Onami, "The Sperm TRP-3 Channel Mediates the Onset of a Ca2+ Wave in the Fertilized C. elegans Oocyte", Cell Reports, doi: 10.1016/j.celrep.2016.03.040
発表者
理化学研究所
生命システム研究センター 細胞動態計測コア 発生動態研究チーム
チームリーダー 大浪 修一 (おおなみ しゅういち)
研究員 髙山 順 (たかやま じゅん)
 大浪 修一
大浪 修一
 髙山 順
髙山 順
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
科学技術振興機構 広報課
Tel: 03-5214-8404 / Fax: 03-5214-8432
jstkoho [at] jst.go.jp(※[at]は@に置き換えてください。)
JST事業に関すること
舘澤 博子(たてさわ ひろこ)
科学技術振興機構 バイオサイエンスデータベースセンター(NBDC)
Tel: 03-5214-8491 / Fax: 03-5214-8470
nbdc-kikaku [at] jst.go.jp(※[at]は@に置き換えてください。)
補足説明
- 1.線虫 C. elegans
遺伝学のモデル生物として広く使用されている自活性の線虫。学名を Caenorhabditis elegansという。ほとんどが雌雄同体で、まれに雄が生まれる。特長として、体が無色透明であることから生きたまま細胞の中を顕微鏡で観察できることが挙げられる。また、動物として初めて全ゲノム配列が解読されたことや、発生時の細胞分裂パターン(細胞系譜)がすべて分かっていること、神経回路の接続パターンがすべて明らかにされていることなど基礎的な情報が整備されている。 - 2.カルシウム透過性チャネル
細胞の膜に存在し、ポアと呼ばれる穴を通じてイオンを透過するタンパク質をチャネルと呼ぶ。カルシウムイオン(Ca2+)を透過するチャネルをカルシウム透過性チャネルと呼ぶ。 - 3.受精カルシウム波
受精時に卵子内に生じる、カルシウムイオン濃度変化が伝播していく現象のこと。1978年米国の研究グループがメダカを用いた実験により初めて観察した。その後、有性生殖を行う動物および植物において広く観察されることが分かった。受精カルシウム波は、種によってその時空間パターンが異なり、一過性の進行波の形状をとるものや、振動を示すものなど多様なものが知られている。 - 4.全能性
多細胞生物の体は様々な種類の細胞から成り立っているが、そのすべての種類に分化しうる性質を全能性と呼ぶ。例えば動物のすべての細胞は受精卵に由来するため、受精卵は全能性を有していると言える。 - 5.カルシウム貯蔵庫
通常、細胞質のカルシウムイオン濃度は、数十~100ナノモーラー(nM、1nMは10億分の1M)程度と非常に低く保たれている。また、細胞には小胞体と呼ばれる細胞内小器官が存在し、小胞体の中のカルシウムイオン濃度は数百マイクロモーラー(µM、1µMは100万分の1M)~数ミリモーラー(mM)程度と考えられている。このように高濃度のカルシウムイオンを保持した細胞内小器官をカルシウム貯蔵庫と呼ぶ。 - 6.蛍光カルシウム指示薬
細胞内のカルシウム濃度に応じて、その蛍光強度または2色の蛍光強度の比を変化させる合成化合物のこと。これを用いることで、細胞内のカルシウム濃度の空間的な変化を観察できる。 - 7.スピニングディスク共焦点顕微鏡
共焦点顕微鏡は、ピンホールを利用することで立体的なサンプルの特定の平面のみを観察することを可能にした顕微鏡。スピニングディスク共焦点顕微鏡は、ニポウディスクと呼ばれる共焦点ピンホールを多数並べた回転する円盤を用いることで、各点の同時観察を可能にした共焦点顕微鏡の1種である。高速なイメージングができ、細胞へのダメージも少ないという特長がある。
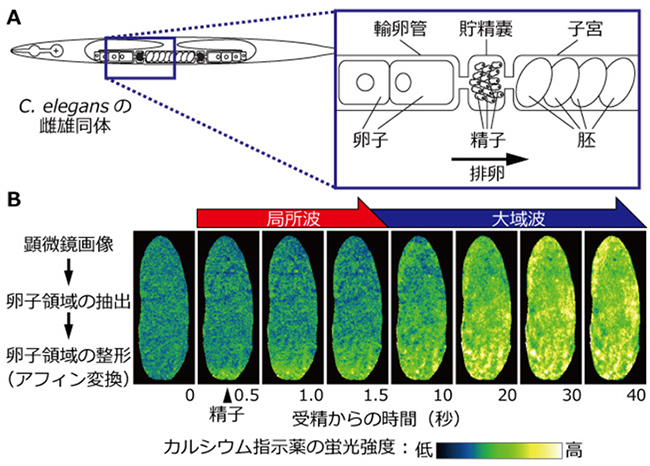
図1 C. elegansの受精カルシウム波の可視化
A:C. elegansの雌雄同体の全体構造と、受精の場である貯精嚢付近の模式図。輸卵管に並んだ卵子は約20分ごとに貯精嚢へ排卵される。貯精嚢には精子が複数存在し、この中の1個の精子が受精する。その後、受精した卵子は子宮へと運ばれ胚となる。
B:観察画像そのものでは、視野内を卵子が移動するため、カルシウム濃度変化の空間的特徴が捉えにくい。そのため画像処理技術を用いて顕微鏡画像から卵子領域を抽出し、一定の形に整形することで、カルシウム濃度変化がどのように広がっていくかを可視化した。その結果、受精直後に精子侵入点付近(画像では▲で示される卵子の真下の部分)でみられる急激で局所的なカルシウム濃度上昇(局所波)と、それに引き続いて起こる卵子全体を伝播する波(大域波)の二相性の波を形成することが分かった。
YouTube:線虫C. elegansの野生型の受精カルシウム波の時空間動態(動画)
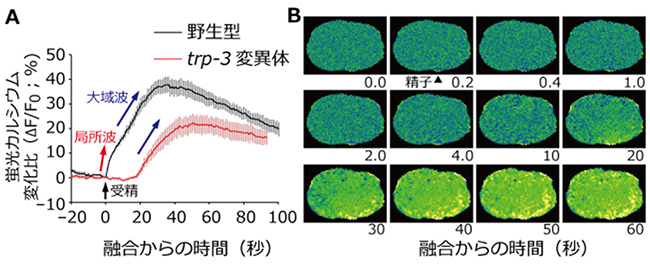
図2 trp-3変異体の受精カルシウム波
A:野生型およびtrp-3変異体の受精時の卵子が示すカルシウム濃度変化のグラフ。縦軸は、卵子全体のカルシウム蛍光強度の平均値を受精直前を基準として表したもの。trp-3変異体では受精直後のカルシウム濃度上昇がなく、約20秒経過してから応答が始まる。
B:trp-3変異体の受精時の卵子のカルシウム濃度変化を表す画像。受精から20秒ほど経過した後に、精子侵入点付近からカルシウム蛍光強度が上昇することが分かる。図2Bは図1Bと同様の画像処理を施したものである。野生型は図1Bを参照。
YouTube:線虫C. elegansのtrp-3変異体では局所カルシウム波が見られない。(動画)
YouTube:線虫C. elegansのtrp-3変異体では大域カルシウム波が遅れて生じる。(動画)
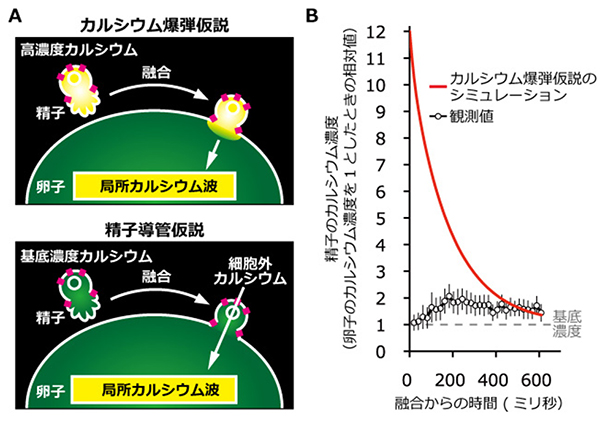
図3 C. elegansの精子カルシウム濃度変化の観測値とカルシウム爆弾仮説のシミュレーション
A:カルシウム爆弾仮説と精子導管仮説の比較の模式図。赤い四角はカルシウム透過性チャネルを示す。上のカルシウム爆弾仮説では、受精前の精子が高濃度のカルシウムをあらかじめ溜め込んでおり、融合によってこれを卵子内に放出し、これが局所波として現れると考える。下の精子導管仮説では、受精前の精子は卵子と同等の基底濃度のカルシウムを持っているが、融合により受精卵の細胞膜の一部となったカルシウム透過性チャネルが細胞外からカルシウムを流入することで、局所波を形成すると考える。両仮説の違いは、受精直後の精子の細胞内カルシウム濃度を測定することで区別できる。
B:受精直後の精子細胞質のカルシウム濃度の観測値と、カルシウム爆弾仮説に基づくシミュレーションの値の比較のグラフ。観測からは基底カルシウム濃度(1として示した)から徐々に上昇していくことが分かる。一方、カルシウム爆弾仮説では高濃度から減少することが予測される。カルシウム爆弾仮説は観測値と一致しなかった。
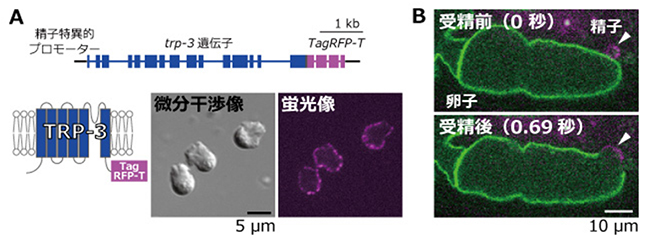
図4 精子のTRP-3チャネルを赤色、卵子の細胞膜を緑色に標識したC. elegansの受精
A:模式図は、TRP-3チャネルの精子細胞内の局在を調べるための融合遺伝子の構造。画像は、TRP-3チャネルにTagRFP-Tという赤色の蛍光タンパク質を融合させたものを、C. elegansの精子(微分干渉像に示されたアメーバ状の細胞)に発現させたもの。精子の細胞膜の蛍光シグナルからTRP-3チャネルが細胞膜に局在することが分かる。
B:卵子と精子の細胞膜が融合する瞬間の可視化。卵子は、細胞膜の脂質成分をGFP::PHという蛍光タンパク質で緑色に蛍光標識したものを利用した。精子はAで作製したものを利用した。受精の瞬間にお互いの細胞膜が融合することが分かる。
YouTube:線虫C. elegansにおける卵子と精子の細胞膜が融合する様子(動画)