要旨
理化学研究所(理研)脳科学総合研究センター行動神経生理学研究チームの村山正宜チームリーダー、名古屋大学環境医学研究所の山中章弘教授、東京大学大学院薬学系研究科の松木則夫教授(研究当時)らの共同研究グループ※は、睡眠不足でも大脳新皮質を再活性化することで記憶力が向上することを発見しました。
睡眠には、起きている間の知覚体験を記憶として定着させる機能があります。感覚情報などの外部からの入力が少ない睡眠時の脳内において、内因的な情報により知覚記憶が定着すると考えられています。しかし、具体的にどの脳回路が知覚記憶の定着に関与するかは不明です。大脳新皮質内の第二運動野(M2)[1]という高次な領域は、第一体性感覚野(S1)[2]という低次な領域と互いにつながり「トップダウン回路[3]」を形成しています。村山チームリーダーらは2015年、M2からS1への「トップダウン入力」がマウスの皮膚感覚の正常な知覚に関与することを明らかにしました。そこで今回、共同研究グループは、トップダウン回路が知覚記憶の定着に関与する可能性を探りました。
共同研究グループは、マウスにおいて、知覚学習直後の深い眠り(ノンレム睡眠[4])時にトップダウン入力を抑制すると、知覚記憶の定着が妨げられることを見出しました。また、M2とS1の神経細胞の活動を記録すると、学習時とノンレム睡眠中に活動が上昇しており、睡眠中に再活性化されることが分かりました。さらに、M2とS1の神経細胞群の活動の同期性は、ノンレム睡眠時に上昇していました。そこで、学習後のノンレム睡眠時にマウス大脳新皮質のM2とS1を同期して刺激したところ、マウスは学習した知覚記憶をより長く保持することが分かりました。
ヒトや実験動物において、長時間にわたって眠らせない「断眠」を行うと、記憶の定着が阻害されることが知られています。ところが、学習後のマウスを断眠させながら大脳新皮質のM2とS1を同期して刺激した場合では、通常の睡眠をとったマウスと比べても、より長い間知覚記憶を保持しました。
共同研究グループは、学習直後の断眠時に大脳新皮質を再活性化させることで、睡眠不足でも知覚記憶を向上できることを証明しました。知覚記憶の定着は睡眠に特有な機能ではなく、睡眠不足による記憶力の低下は脳刺激によって補えることが分かりました。
今後、マウスにおける大脳新皮質の刺激パターンをさらに臨床に適用できるよう改良することで、睡眠障害による記憶障害の治療方法の開発に応用できると期待できます。
本研究は、米国の科学雑誌『Science』に掲載されるのに先立ち、オンライン速報版(5月26日付け:日本時間5月27日)に掲載されます。
※共同研究グループ
理化学研究所 脳科学総合研究センター
行動神経生理学研究チーム
チームリーダー 村山 正宜(むらやま まさのり)
客員研究員(日本学術振興会特別研究員PD) 宮本 大祐(みやもと だいすけ)
リサーチアソシエイト 平井 大地(ひらい だいち)
研究員 鈴木 崇之(すずき たかゆき)
テクニカルスタッフⅠ 小田川 摩耶(おだがわ まや)
テクニカルスタッフⅠ 松原 智恵(まつばら ちえ)
神経回路・行動生理学研究チーム
チームリーダー トーマス・ジェイ・マクヒュー(Thomas J McHugh)
テクニカルスタッフⅠ ロマン・ボーリンガー(Roman Boehringer)
研究員 チナカルパン・アダイカン(Chinnakkaruppan Adaikkan)
脳回路機能理論研究チーム
チームリーダー 深井 朋樹(ふかい ともき)
研究員 チ・チュン・アラン・ファン(Chi Chung Alan Fung)
名古屋大学 環境医学研究所 ストレス受容・応答研究部門 神経系分野II
教授 山中 章弘 (やまなか あきひろ)
特任助教(研究当時) 犬束 歩(いぬつか あゆむ)
東京大学大学院 薬学系研究科 薬品作用学
教授(研究当時) 松木 則夫(まつき のりお)(現 名誉教授)
背景
睡眠はなぜ必要なのでしょうか?1日の多くの時間を占める睡眠の機能は、意外にもあまりよく分かっていません。現代人の多くは睡眠不足と言われていますが、わずかな睡眠不足でも脳機能に影響を及ぼすことは実感するところです。睡眠の機能を理解し、睡眠不足に対する適切な対処法を探ることは、健康な生活や生活の質の向上のために重要な課題です。
これまで知られている睡眠の機能の1つとして「記憶の定着作用」があります。例えば、起きている間に体験したさまざまな感覚は「知覚記憶」として睡眠中に定着します。感覚情報の知覚は体の各部位から脳の低次な領域、そしてより高次な領域へと、情報が伝わることで起こります。ところが睡眠時には、感覚情報を知覚する機能が低下していて、新たな感覚情報はあまり入ってきません。そのため知覚記憶の定着には、脳のより高次な領域から低次な領域へと、逆に情報が伝わる「トップダウン回路」を介した内因的な情報処理が重要であると考えられてきました。しかし、このような感覚情報処理に関与する脳回路が特定されなかったことから、実際にトップダウン回路を操作した場合、知覚記憶の定着が向上したり低下したりするのかは不明でした。
村山チームリーダーらは2015年、触覚の知覚に関わる大脳新皮質のトップダウン回路を発見しています注)。大脳新皮質の第二運動野(M2)という領域から第一体性感覚野(S1)という領域にトップダウン回路が形成されており、マウスのM2からS1へのトップダウン入力を抑制すると、皮膚感覚の正常な知覚が阻害されることを見出しました。そこで今回、共同研究グループは、M2やS1における神経活動の操作が知覚記憶の定着に与える影響を調べることにしました。
注)2015年5月22日プレスリリース「“感じる脳”のメカニズムを解明」
研究手法と成果
共同研究グループはまず、触覚に関する知覚記憶の定着を調べるためのマウスの行動課題を確立しました(図1)。1日目、マウスを10分間、ツルツルの床の上で探索させたのち、元のケージに戻します。床の左右には探索用のオブジェクトを置いておきます。2日目にマウスを、半分がツルツル、半分がデコボコの床に置きます。すると、マウスはなじみのない新しい環境を好んで探索する性質(選好性)があるので、ツルツルの床面に置かれたオブジェクトよりも、初めて体験するデコボコの床面に置かれたオブジェクトをより長い時間探索するようになります。2日目に見られる、一方の質感の床面における滞在時間の偏りは、前日に経験した床面の質感を記憶し、その記憶に基づいた新しい質感の床面への選好性を示していると考えられ、知覚記憶の指標として用いました。
この行動課題に睡眠が必要であるかどうかを調べるため、マウスが動かなくなる睡眠中にケージを揺らして睡眠を阻害し、断眠処置を行いました。マウスが学習した直後の1時間を断眠すると、2日目における新規な床面への選好性が低下しました。一方、学習の6時間後の1時間を断眠すると記憶成績は影響を受けませんでした。これより、学習直後の睡眠が知覚記憶の定着に必要であることが分かりました。
マウスの脳状態は、少なくとも3つに分類できます。「覚醒状態」、深い眠りである「ノンレム睡眠」、浅い眠りである「レム睡眠[5]」です。そこで、マウスの脳波と筋電位からリアルタイムで脳状態を判定し、特定の脳状態の間だけ、M2からS1への投射を光遺伝学的手法[6]により抑制しました(図2)。学習直後(0~1時間)のノンレム睡眠時に光照射によりトップダウン回路を抑制すると、知覚記憶の定着は阻害されましたが、学習してから6~7時間後のノンレム睡眠時の光照射は、記憶の定着に影響しませんでした(図2D)。一方、学習直後の覚醒時に光照射によって同じ回路を抑制しても、記憶の定着には影響はありませんでした。これより、学習直後のノンレム睡眠時におけるトップダウン入力が記憶の定着に必要であることが分かりました。
記憶の定着がどのような神経活動によって担われているかを調べるために、M2とS1に電極を刺入し、各領域の局所フィールド電位[7]を記録しました(図3)。ノンレム睡眠時では、デルタ帯域(0.5~4Hz)のゆっくりとした脳波がM2とS1とで観察され、同期していました。この2領域での脳波をGranger 因果性解析[8]すると、実際にM2からS1へ情報が流れていくことを見出しました。光照射でM2からS1へのトップダウン入力を抑制すると、この情報の流れが抑制されました。
次に、それぞれの領域での単一神経細胞レベルの神経活動を記録しました。するとM2とS1の両方において、学習時に活性化した神経細胞は、学習後のノンレム睡眠時においても活性化していました。これを「再活性化」と呼びます。光照射によってM2からS1へのトップダウン入力を抑制すると、M2における再活性化は阻害されませんでしたが、S1における再活性化が阻害されました。これまでの研究で、記憶の定着には神経活動の再活性化が重要だと考えられてきましたが、実際にどの脳領域が再活性化を誘発しているのか、全く分かっていませんでした。これらの結果は、感覚記憶の定着に不可欠な感覚野における再活性化には、皮質間トップダウン入力が必要であることを示しました。
睡眠時にトップダウン回路を抑制すると記憶の定着が阻害されましたが、では逆に、この回路を活性化したらどうなるでしょうか。大脳新皮質を刺激するために、大脳新皮質にチャネルロドプシン(ChR2)[9]を発現した遺伝子改変マウス[10]を用いました。学習後のノンレム睡眠時にM2とS1を、2Hzのリズムで同期させた光を用いて刺激(同期光刺激)しました(図4)。光刺激を行わない通常のマウスでは、学習してから4日後に床面認識課題を行うと、1日目の知覚記憶を忘れており、床面の選好性を示しませんでした。ところが同期光刺激したマウスでは、4日経っても記憶を保持していました。一方で、光を同期させずにM2とS1を刺激すると、翌日にテストした場合でも、記憶の定着が阻害されました。この結果は、特定の脳刺激法により、記憶の定着をコントロールすることが可能であることを示します。
最後に、断眠されたマウスでも同期光刺激によって記憶の定着が可能かを調べました。断眠させない通常のマウスにおける正常なノンレム睡眠パターンに合わせた光刺激パターンで、断眠中のマウスのM2とS1に同期光刺激を与えたところ、このマウスは、2日目にテストした場合も4日後にテストした場合も、知覚記憶を保持していました。このことから睡眠不足の場合でも、大脳新皮質を適切に刺激すれば、記憶を維持、向上させることが可能であることが分かりました。
今後の期待
共同研究グループは、知覚記憶の定着には、学習直後のノンレム睡眠時、すなわち寝入り時におけるトップダウン入力が必要であることを示しました。また、記憶の定着に関連する神経活動の再活性化は、このトップダウン入力が関連することを示しました。さらに、睡眠不足の状態でも、適切なタイミングで大脳新皮質を刺激することで、知覚記憶を向上できることを示しました。質の良い睡眠は心と体を正常に保つために重要ですが、知覚記憶の定着に関しては、睡眠が必要ではない可能性を示す、驚くべき結果であるといえます。一般的に、マウスは約12時間の睡眠をとります。トップダウン回路の光抑制や光刺激は合計30分間行いました。たった4%の睡眠時間が記憶力の向上や低下に関与していたことは驚きです。
M2とS1は皮膚感覚の知覚情報に関連した感覚野ですが、他の感覚野(視覚野、聴覚野等)でも似たような記憶の定着メカニズムがある可能性があります。今回、光刺激を行った感覚野を含む大脳新皮質は脳表面に位置します。近年臨床で用いられ始めている経頭蓋(けいとうがい)磁気刺激[11]や経頭蓋直流刺激[12]によって、ヒトにおいても刺激を与えることが可能です。また、睡眠障害を持つ患者などでは記憶力の低下が起きているかも知れません。今回見出したマウスの記憶の定着を向上させる刺激パターンを、臨床に適用・改良することで、そのような患者の治療法を開発できる可能性があります。また、健常なヒトに応用した場合は知覚記憶の向上や、高齢者における知覚記憶の維持などが可能になるかもしれません。
原論文情報
- Daisuke Miyamoto, Daichi Hirai, Chi Chung Alan Fung, Ayumu Inutsuka, Maya Odagawa, Takayuki Suzuki, Roman Boehringer, Chinnakkaruppan Adaikkan, Chie Matsubara, Norio Matsuki, Tomoki Fukai, Thomas J McHugh, Akihiro Yamanaka & Masanori Murayama, "Top-Down Cortical Input during NREM Sleep Consolidates Perceptual Memory", Science, doi: 10.1126/science.aaf0902
発表者
理化学研究所
脳科学総合研究センター 行動神経生理学研究チーム
チームリーダー 村山 正宜(むらやま まさのり)
名古屋大学 環境医学研究所 ストレス受容・応答研究部門 神経系分野II
教授 山中 章弘(やまなか あきひろ)
東京大学大学院 薬学系研究科 薬品作用学
教授(研究当時) 松木 則夫(まつき のりお)(現 名誉教授)
 村山 正宜
村山 正宜
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
名古屋大学総務部広報渉外課
Tel: 052-789-2699 / Fax: 052-788-6272
kouho [at] adm.nagoya-u.ac.jp(※[at]は@に置き換えてください。)
補足説明
- 1.第二運動野(M2)
大脳新皮質の前頭野に位置し、動物行動の計画や実行、報酬や記憶に関連した活動を示す高次脳領域。 - 2.第一体性感覚野(S1)
大脳新皮質の頭頂葉に位置し、皮膚感覚や深部感覚をつかさどる低次脳領域。S1内の各所と体部位には対応関係がある。 - 3.トップダウン回路
外界の情報は受容器を介して、中枢神経系を通って脳へ伝わる。脳内で中枢神経系から最初に情報を受け取る領域を低次脳領域と呼び、低次脳領域からさらに情報を受け取ってより複雑な処理をする領域を、高次脳領域と呼ぶ。低次領域から高次領域への経路をボトムアップ回路と呼ぶが、逆に高次脳領域から低次脳領域に情報が伝わるような経路をトップダウン回路と呼ぶ。 - 4.ノンレム睡眠
深い睡眠で、脳波は大きな低周波のデルタ波を生じる。 - 5.レム睡眠
浅い睡眠で、脳波は小さくて覚醒状態時に近い。 - 6.光遺伝学的手法
神経回路機能を光と遺伝子操作を使って調べる手法。ミリ秒(0.001秒)単位の時間精度を持った制御を特徴とする。神経回路を抑制するために、抑制性の光感受性イオンポンプ(ArchT)の遺伝子配列が組み込まれたアデノ随伴ウイルス(AAV)を脳内に注入することで、神経細胞にArchTを発現させた。その神経細胞に光を照射すると、光感受性イオンポンプが活性化し、細胞内から細胞外にH+が排出され、結果的に神経細胞の活動は抑制される。 - 7.局所フィールド電位
記録電極周囲の神経細胞集団全体の活動を調べる方法。局所フィールド電位は、集団活動のオン・オフのリズムと関連している。 - 8.Granger 因果性解析
ノーベル経済学賞受賞者Granger博士による、XからYの予測を試みる解析方法。経時変化する2つの変数XとY(例: 気温とビールの消費量)があるとき、変数Y(例: ビールの消費量)の今の値が、Y自身の過去の値だけから予測するよりも、Xの過去の値(気温)を考慮した方がより精度良く予測できる場合、「変数Xは変数Yに対してGranger因果性がある」といい、XがYに対して影響を及ぼしていることが分かる。この解析法は近年、脳波等の脳信号の解析に応用されている。 - 9.チャネルロドプシン(ChR2)
藻類のクラミドモナスの眼点から同定された、光活性型非選択的陽イオンチャネルタンパク質。青色光によって活性化すると、チャネルが開き、プラスに帯電した陽イオンが細胞外から細胞内に流入するため、このチャネルを発現した神経細胞は活性化される。 - 10.遺伝子改変マウス
外来性の遺伝子を持つように遺伝子改変をしたマウス。本研究で用いたThy1-ChR2マウスでは、大脳新皮質の神経細胞に興奮性の光感受性イオンチャネルであるChR2を発現している。その神経細胞に青色光を照射すると、光感受性イオンチャネルが活性化し、細胞外から細胞内に陽イオンが流入し、結果的に神経細胞の活動は活性化される。 - 11.経頭蓋磁気刺激
頭皮に置いた電極から局所的な強い磁場をかけることで、脳内の神経細胞を活性化する方法。 - 12.経頭蓋直流刺激
頭皮の上に置いた電極で微弱な直流電気を通電する手法
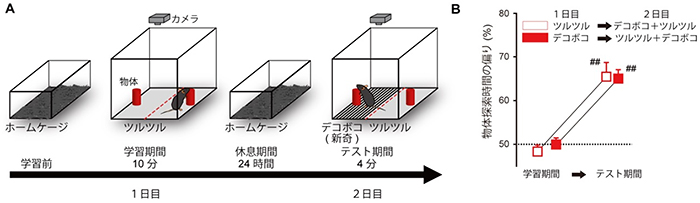
図1 床面認識課題
- A.同じ物体を左右の床に置き、マウスがこの物体(オブジェクト)を探索する時間を計測する。1日目は、ツルツル、またはデコボコの一方のみを床面とする。2日目は、半分がツルツル、もう半分がデコボコの床面を用いる。
- B.マウスの物体探索時間。学習期間(1日目)では、左右同一の質感の床では物体探索時間に偏りは見られない。テスト期間(2日目)では、新奇の質感の床面上にある物体を探索する時間が増える。
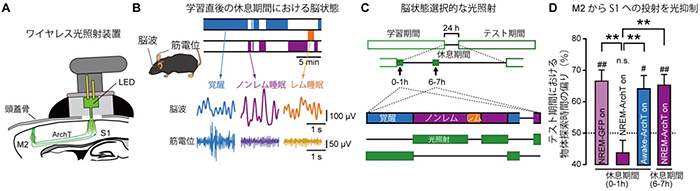
図2 学習後の睡眠時における光抑制
- A.第二運動野(M2)と第一体性感覚野(S1)への神経投射を特異的に抑制するための光照射装置。M2には光感受性抑制性ポンプであるArchTを遺伝学的手法によって導入し、光照射装置でM2からS1へのトップダウン入力を抑制した。
- B.マウスの脳状態を、脳波と筋電位より同定した。
- C.脳状態選択的に光照射を行った。光照射の合計時間は30分とした。
- D.M2からS1への神経投射を、学習直後(0~1時間)のノンレム睡眠時に光照射により抑制した場合、2日目のテスト期間での物体探索時間の偏りが減少した。しかし、学習後6~7時間後のノンレム睡眠時に光照射した場合では、物体探索時間の偏りに変化はなかった。
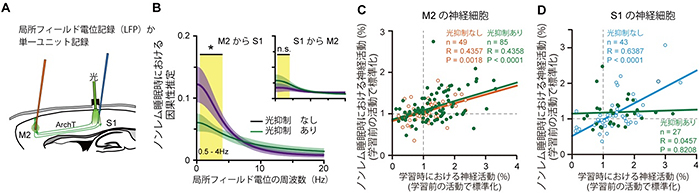
図3 光遺伝学を用いたトップダウン回路抑制による神経活動への影響
- A.第二運動野(M2)と第一体性感覚野(S1)から、局所フィールド電位記録または単一ユニット記録を行った。M2からS1へのトップダウン入力を抑制した。
- B.光抑制あり・なしにおける M2-S1間のGranger因果性解析。光抑制によりM2とS1間の情報の流れは阻害されていた。
- C.光抑制によるM2神経細胞の再活性化の影響。各丸は各単一神経細胞の活動を表す。光抑制のあり・なし、どちらの条件でも相関の傾きに変化がないことから、光抑制によって、M2神経細胞の再活性化は影響されないことが分かる。
- D.光抑制によるS1神経細胞の再活性化の影響。光抑制ありの場合は、相関がなくなることから、再活性化が抑制されていることが分かる。
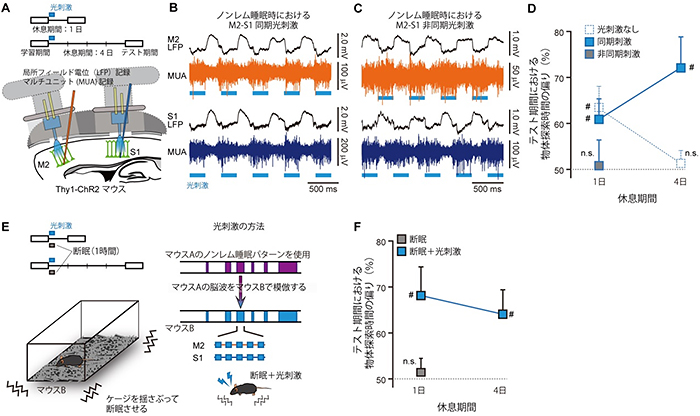
図4 光刺激による知覚記憶の定着への影響
- A.光刺激装置。Thy1-ChR2遺伝子改変マウスを用いた。光刺激を学習直後のノンレム睡眠時に行った。
- B.M2とS1とを同期刺激した際の神経活動。
- C.M2とS1とを非同期で刺激した際の神経活動。
- D.同期/非同期刺激した場合の物体探索時間の偏り。
- E.M2とS1とを同期光刺激しながらマウスを断眠させた。光刺激は、別のマウスAにおける正常なノンレム睡眠パターンを型としてマウスBに行った。
- F.断眠のみのマウスでは記憶の定着が阻害されるが、これに光刺激を加えたマウスでは、記憶の定着が向上した。