要旨
理化学研究所(理研)環境資源科学研究センター合成ゲノミクス研究グループの岡義人基礎科学特別研究員(研究当時)、吉積毅研究員(研究当時)、松井南グループディレクター、カリフォルニア大学ロサンゼルス校のチェンタオ・リン教授らの国際共同研究グループ※は、植物の細胞核内に存在する青色光受容体のクリプトクロムが青色光を受容すると「二量体化」することで活性化されること、BIC1タンパク質がクリプトクロムの青色光依存的二量体化を阻害することによりクリプトクロムの活性を調節していることを明らかにしました。
植物にとって光は、光合成によるエネルギー源となるだけでなく、周囲の光環境を知るための情報源としても重要な役割を担っています。植物は情報としての光を感知するために、いくつかの光受容体を進化させてきました。中でもクリプトクロムは、脱黄化[1]、花芽形成、避陰反応[2]など数多くの植物の青色光応答を制御する重要な光受容体です。クリプトクロムは青色光を受容して活性化されると、さまざまな遺伝子の発現を調節して植物の青色光応答を誘導します。しかし、青色光によるクリプトクロムの活性化の分子機構はほとんど明らかになっていませんでした。さらに、クリプトクロムの脱感作[3]機構に関しても、一部のクリプトクロム分子種が青色光を受容すると分解されることが知られているのみで、全ての分子種に共通する機構は見つかっていませんでした。
国際共同研究グループは、クリプトクロムの新たな脱感作機構を明らかにするために、シロイヌナズナの完全長cDNA高発現系統「ナズナFOXライン[4]」を用いて、青色光応答が低下した変異体のスクリーニングを行いました。その結果、クリプトクロム情報伝達に抑制的に働く「BIC1(Blue light Inhibitor of Cryptochrome 1)遺伝子」を発見しました。また、BIC1はクリプトクロムと結合し、クリプトクロムのリン酸化やクリプトクロムが制御する種々の遺伝子発現制御など、クリプトクロムの情報伝達に関わる反応を全て阻害しました。また、クリプトクロムが青色光に応じて、二量体を形成することが明らかになりました。さらに、BIC1はクリプトクロムの青色光依存的な二量体化形成を阻害しました。これらの結果は、青色光依存的な二量体化形成がクリプトクロムの初期反応を担う重要な過程であり、BIC1はクリプトクロムの二量体化を阻害することで、それに引き続くクリプトクロムのリン酸化などの一連の反応を抑制していることを示しています。
クリプトクロムはさまざまな農業上の重要形質を制御しているため、本研究の成果は今後、作物量のバイオマス増収などにつながると期待できます。
本研究は、米国の科学雑誌『Science』に掲載されるのに先立ち、オンライン速報版(10月20日付け:日本時間10月21日)に掲載されます。
※国際共同研究グループ
理化学研究所 環境資源科学研究センター 合成ゲノミクス研究グループ
基礎科学特別研究員(研究当時) 岡 義人(おか よしと)(現 合成ゲノミクス研究グループ 客員研究員、中国・福建農林大学教授)
研究員(研究当時) 吉積 毅(よしづみ たかし)(現 酵素研究チーム上級研究員)
グループディレクター 松井 南(まつい みなみ)
カリフォルニア大学ロサンゼルス校 分子細胞発生生物学部
教授 チェンタオ・リン(Chentao Lin)
全南大学校 バイオテクノロジー・クムホ ライフサイエンス学部
教授 ジョンイル・キム(Jeong-Il Kim)
背景
植物にとって光は、光合成によるエネルギー源となるだけでなく、周囲の光環境を知るための情報源としても重要な役割を担っています。そのため、植物は周囲の光環境を察知して光合成を効率よく行うために、形態や光受容などを最適化しています。中でも、細胞核内で主に機能するクリプトクロムは、脱黄化、花芽形成、避陰反応など農業で重要な数多くの生長・発達の過程を制御する重要な光受容体です(図1)。その情報伝達機構が解明されれば、将来の農業への応用につながると期待されています。
クリプトクロムは青色光を受容することで活性化され、間接的もしくは直接的に転写因子[5]の活性を調節することで、種々の遺伝子の発現を制御します。活性化したクリプトクロムは、タンパク質分解に関わるCOP1/SPA1複合体と結合するとCOP1/SPA1複合体を不活性化し、HY5などのCOP1/SPA1複合体の標的転写因子の蓄積を促すことで、間接的に転写因子の活性を調節します。また、CIB1などの転写因子との直接的な結合により、これら転写因子の活性を調節します。このように、活性型クリプトクロムが青色光情報を下流の因子へ伝達する分子機構は比較的よく理解されていますが、青色光受容によりクリプトクロムがどのように活性化されるのかは、これまで全く明らかにされていませんでした。
また、他の光受容体で一般的にみられるような脱感作機構に関しては、シロイヌナズナに存在する二つのクリプトクロム分子種CRY1とCRY2のうち、CRY2が青色光を受容して活性型になると速やかに分解されることが知られています。しかし、CRY1とCRY2に共通するクリプトクロムの脱感作機構は明らかにされていませんでした。
注)光生物特有の表記法について
光生物分野では大文字/小文字と斜体/非斜体を組み合わせることで特定の種類の遺伝子、タンパク質であることを表記する。本リリースでは、cry1cry2(小文字斜体)は遺伝子変異、CRY1CRY2(大文字斜体)は遺伝子、CRY1CRY2(大文字正体)はクリプトクロムタンパク質である。
研究手法と成果
最初に、国際共同研究グループは、クリプトクロムの脱感作機構を解明するため、理研が開発したシロイヌナズナのFOX hunting system[6]という機能付加型変異体ライブラリーであるナズナFOXラインを用いて、青色光応答が低下した変異体について遺伝学的スクリーニングを行いました。
その結果、クリプトクロム情報伝達に抑制的に働く「BIC1 (Blue light Inhibitor of Cryptochrome 1)遺伝子」を過剰に発現するbic1D変異体を単離しました。bic1D変異体は、cry1cry2二重変異体と同様に青色光応答が低下し、青色光条件下で胚軸(発芽した苗の茎の部分)が徒長[7]しました。また、シロイヌナズナのBIC1相同遺伝子であるBIC2と緑色蛍光タンパク質のGFPの融合遺伝子(BIC2-GFP)の過剰発現体の光応答を調べたところ、bic1D変異体と同様に青色光応答が低下しました。
一方、bic1変異体やbic2変異体は、はっきりとした表現型を示さないものの、bic1bic2二重変異体はbic1D変異体とは逆に、青色光に対して高い感受性を示しました。さらに、bic1bic2cry1cry2四重変異体がcry1cry2二重変異体と同様の表現型を示し青色光応答が低下したことから、BIC1とBIC2はクリプトクロムの下流でクリプトクロムの情報伝達を抑制することで、脱感作を促していることが明らかになりました。
CRY1とCRY2は、いずれも青色光に応答してリン酸化されます。クリプトクロムの機能とリン酸化との間には密接な相関があることから、リン酸化がクリプトクロムの情報伝達において重要な役割を担っていると考えられます。本研究で、bic1D変異体やBIC2-GFPの過剰発現体においては、CRY1とCRY2のリン酸化は全く検出されませんでした。さらに、酵母ツーハイブリッド法[8]や、ヒト胎児由来腎臓細胞であるHEK293細胞、もしくは植物細胞の免疫沈降実験[9]などにより、CRY2とBIC1が結合することが明らかになりました。これらの結果は、BIC1がクリプトクロムに直接結合することにより、リン酸化などの情報伝達を抑制することを示しています。
クリプトクロムの二量体化は、クリプトクロムのリン酸化やそれに引き続く種々の遺伝子の発現など、クリプトクロムの情報伝達に必須であることが知られています。上述の通り、BIC1はクリプトクロムと直接結合し、リン酸化を阻害することから、BIC1や同様にBIC2のクリプトクロムとの直接的な結合が、クリプトクロムの二量体化を阻害する可能性があります。一方、CRY1とCRY2はいずれも、植物体内で青色光の有無に関わらず二量体化することが示されていました。しかし過去の報告では、過剰に発現したエピトープタグ[10]融合クリプトクロムを用いて免疫沈降実験を行うことで、内在性のクリプトクロムを検出していました。そのため、CRY1とCRY2の量比の違いにより本来のクリプトクロムの二量体化の青色光依存性が見かけ上損なわれていた可能性がありました。
そこで、国際共同研究グループは、クリプトクロムの青色光依存的な二量体化を検証するために、HEK293細胞、もしくは植物体内で異なるエピトープタグと融合させた二種類のCRY2融合タンパク質を共発現させた植物細胞を用いて免疫沈降実験を行いました。
その結果、HEK293細胞、植物細胞いずれにおいても、CRY2タンパク質は青色光に応じて二量体を形成することが明らかになりました。CRY2の青色光依存的二量体化は、酵母ツーハイブリッド法や植物体内での分割蛍光タンパク質再構成法[11]でも確認されました。また、同様の方法でBIC1が植物細胞、HEK293細胞、酵母細胞において、青色光依存的CRY2の二量体化を阻害することが示されました。さらに、HEK293細胞を用いた免疫沈降実験により、BIC1がCRY2とSPA1、あるいはCIB1との青色光依存的な結合も阻害することが明らかになりました。
以上の結果と、二量体化がクリプトクロムのリン酸化などの情報伝達に必要であることを踏まえると、CRY2の青色光依存的な二量体形成がCRY2の青色光応答の初期過程を担う重要な反応であると考えられます。また、BIC1はCRY2の青色光依存的二量体形成を阻害することにより、CRY2のリン酸化や情報伝達因子との結合などのCRY2の一連の情報伝達の過程を抑制していると考えられます(図2)。
今後の期待
本研究によりクリプトクロムの活性化と脱感作の機構が明らかになりました。クリプトクロムが制御する反応には農業上重要な形質も数多く含まれることから、今後はBIC1を利用してクリプトクロムの活性を人為的に操作する系の構築などにより作物バイオマスの増収などにつながると期待できます。
また、クリプトクロムの多量体化やクリプトクロムとCIB1の青色光依存的結合は光遺伝学[12]のツールとしても利用されており、本研究による発見はクリプトクロムを基盤とした光遺伝学技術の改良につながることも期待できます。
さらに、本研究では動物のクリプトクロムが二量体を形成することが明らかになりました。動物のクリプトクロムは体内時計を構成する重要な因子であり、動物のクリプトクロムの二量体形成の生理学的意義の解明は動物の体内時計を支える分子機構の理解に役立つと期待できます。
原論文情報
- Qin Wang, Zecheng Zuo, Xu Wang, Lianfeng Gu, Takeshi Yoshizumi, Zhaohe Yang, Liang Yang, Qing Liu, Wei Liu, Yun-Jeong Han, Jeong-Il Kim, Bin Liu, James A. Wohlschlegel, Minami Matsui, Yoshito Oka, Chentao Lin, "Photoactivation and inactivation mechanisms of Arabidopsis cryptochrome 2", Science, doi: 10.1126/science.aaf9030
発表者
理化学研究所
環境資源科学研究センター バイオマス工学研究部門 合成ゲノミクス研究グループ
基礎科学特別研究員(研究当時) 岡 義人(おか よしと)
研究員(研究当時) 吉積 毅(よしづみ たけし)
グループディレクター 松井 南(まつい みなみ)
カリフォルニア大学ロサンゼルス校 分子細胞発生生物学部
教授 チェンタオ・リン(Chentao Lin)
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.脱黄化
植物を暗所で育てると子葉が黄化する。また、暗所では胚軸は徒長し、子葉は閉じてフックを形成する、いわゆるもやし状の形態を示す。一方、黄化した芽生えを光条件に移すと、胚軸伸長は抑制され、子葉は開いて葉緑体の発達のため緑化する。これら一連の反応を脱黄化と呼ぶ。 - 2.避陰反応
植物は光合成に不利な光環境を避けるため、茎を伸ばしてよりよい光環境に脱出しようとする。これを避陰反応と呼ぶ。 - 3.脱感作
植物に光などの刺激を与え続けることで、初めにあった反応がなくなること。 - 4.ナズナFOXライン
シロイヌナズナの完全長cDNAをそれぞれ高発現する機能付加した、シロイヌナズナ形質転換体の集まりのこと。 - 5.転写因子
転写因子はDNAに特異的に結合するタンパク質の一群であり、DNAの遺伝情報をRNAに転写する過程の調節を行う。 - 6.FOX hunting system
平均化した完全長cDNAをシロイヌナズナにそれぞれ導入して高発現することで機能付加を起こし、優性の形質として遺伝子の機能をみる技術、あるいはそのような遺伝子導入した植物のこと。 - 7.徒長
植物の枝や茎の節の間が伸びること。日照不足になると、草花は日の光を求めて伸びるので徒長しやすい。 - 8.酵母ツーハイブリッド法
酵母細胞を用いて、二種類のタンパク質の結合を調べる手法。 - 9.免疫沈降実験
二種類のタンパク質の結合を、それぞれのタンパク質を同時に発現させた細胞から精製したタンパク質を用いて解析する手法。 - 10.エピトープタグ
免疫沈降や抗体でタンパク質の局在を調べる目的で、既存の抗体が認識する抗原となる配列(エピトープ)をタグとしてタンパク質に結合すること。 - 11.分割蛍光タンパク質再構成法
分割した蛍光タンパク質の再構成による蛍光を指標として、二種類のタンパク質の結合を調べる手法。 - 12.光遺伝学
外部から光により遺伝子発現を制御する新しい手法。遺伝子工学的手法を用いて細胞に特別な光活性型タンパク質を発現させて、光を照射することで、その細胞を活性化させたり抑制させたりする。この手法で簡便に細胞を傷つけることなく遺伝子発現の制御を操作することができるため、近年注目されている技術。
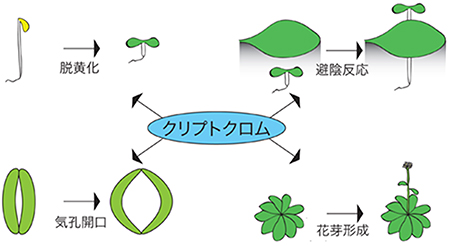
図1 青色光受容体クリプトクロムが制御する生理応答
クリプトクロムは青色光に応答して脱黄化、避陰反応、花芽形成、気孔開口などの重要形質を制御する。
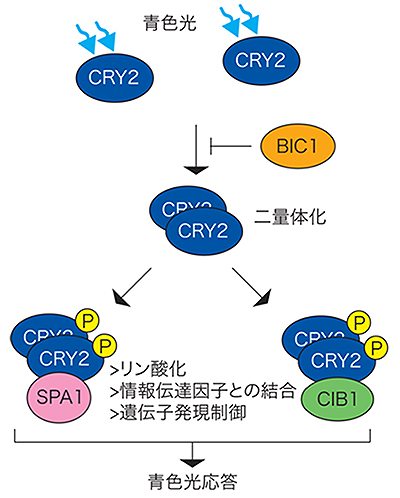
図2 クリプトクロムの青色光応答の制御モデル
クリプトクロム(CRY2)は青色光を受容すると二量体化する。二量体化したCRY2はリン酸化(リン酸は図中のP)などの制御を受け、SPA1やCIB1などとの結合を介して種々の遺伝子の発現を制御し、青色光に応じた形態の変化を誘導する(青色光応答)。BIC1はCRY2と結合することでCRY2の二量体化を阻害し、CRY2の情報伝達を抑制する。