要旨
理化学研究所(理研)統合生命医科学研究センター サイトカイン制御研究チームの久保允人チームリーダー(東京理科大学生命医科学研究所教授)と宮内浩典研究員らの共同研究グループ※は、マウスを用いてインフルエンザウイルスの活性を減退または消失させる抗体(中和抗体)誘導の新しい制御機構を明らかにしました。
インフルエンザウイルスは喉や鼻から体内に侵入して、重篤な肺炎を起こします。鳥インフルエンザなど病原性の高いウイルスが、変異を繰り返してヒトに感染できるようになると、多くのヒトが死亡すると危惧されています。ワクチン接種は、ウイルスが体内へ侵入することを防ぐ抗体を誘導するための有効な手段です。これまで、ワクチン接種による抗体の誘導は、抗体産生の場である「胚中心[1]」と「リンパ濾胞型ヘルパーT細胞(TFH細胞)[2]」の両方が必要と考えられてきました。これは、ウイルスに対して高い結合能(親和性)を持つ抗体が、胚中心でTFH細胞に助けられて作られるためです。したがって、効率よくTFH細胞を活性化することが、効果の高いワクチンの開発につながると考えられていました。
しかし今回、共同研究グループは、季節性インフルエンザウイルスと高病原性鳥インフルエンザウイルスを使って、胚中心やTFH細胞を持たないマウスにおいてワクチン接種によって、中和活性(ウイルスの感染を阻止する作用)の高い「免疫グロブリンG2抗体(IgG2抗体)[3]」が作られることを見出しました。IgG2抗体は、インフルエンザウイルスに対しての親和性は高くありませんが、中和活性が高いためウイルス感染を十分予防できます。また、TFH細胞に代わってインターフェロン-ガンマ(INF-γ)[4]を産生する「Ⅰ型ヘルパーT細胞(TH1細胞)[5]」が、IgG2抗体を誘導することも分かりました。このことから、TH1細胞によって誘導される低親和性の抗体(IgG2抗体など)は、ウイルス抵抗性を付与することが明らかになりました。
本成果により、TH1細胞を活性化することで、低親和性にも関わらず中和活性が高い抗体を産生できることが分かりました。これは、将来起こると予測されるインフルエンザウイルスのパンデミック感染の脅威に対抗する新たな戦術として、効率よくTH1細胞を活性化する新たなワクチンの開発に役立つと考えられます。
本研究は、英国の科学雑誌『Nature Immunology』に掲載されるのに先立ち、オンライン版(10月31日付け:日本時間11月1日)に掲載されます。
背景
インフルエンザウイルスは鼻や喉から体内に侵入し、気道や肺で爆発的に増殖することで重篤な肺炎を引き起こします。鳥インフルエンザウイルスなどの病原性の高いウイルスが、突然変異を繰り返してヒトに感染できるようになると、多くのヒトが死亡すると危惧されています。ワクチン接種は、ウイルスが体内へ侵入することを防ぐ抗体を誘導するための有効な手段です。抗体が産生されるためには、T細胞[6]とB細胞[7]が協調的に働く“抗体産生の場”が必要で、2次リンパ組織[8]の「胚中心」がその場所となっています。
免疫グロブリンG抗体(IgG抗体)は、ウイルスに対する感染防御において、効果の高い抗体として知られています。IgG抗体にはサブクラスがあり、ヒトの場合はIgG1、IgG2、IgG3、IgG4の4種類があります。
これまで抗体産生については、ウイルスタンパク質抗原(ウイルスに含まれる抗原となるタンパク質)に対して作られるIgG1抗体について多く研究されてきました。IgG1抗体産生には、胚中心で抗体産生に特化したリンパ濾胞型ヘルパーT細胞(TFH細胞)とB細胞が結合する必要があることが分かっています。また、胚中心でウイルスに対して高い“結合能(親和性)”を持つ抗体が作られるため、TFH細胞を活性化してより高い親和性を持つ抗体を誘導できるものが、より効果の高いワクチンだと考えられてきました。
一方、IgG2抗体はIgG1抗体と同じく、胚中心でTFH細胞とB細胞が結合して産生されますが、4種類のIgG抗体の中でも、ウイルスの感染を阻止する作用(中和活性)の最も高い抗体であることが知られています。そこで今回、共同研究グループはIgG2抗体に焦点を当てて、抗体産生の研究を進めました。
研究手法と成果
共同研究グループはまず、なぜIgG2抗体の中和活性(ウイルスの感染を阻止する作用)に胚中心やTFH細胞が有効なのかを検証するため、胚中心形成に必要なタンパク質「Bcl-6」に着目しました。Bcl-6は細胞内で遺伝子の転写を制御する分子です。T細胞にBcl-6が発現しないと、T細胞はTFH細胞になることができず、マウスは胚中心を作れません。一方、B細胞にBcl-6が発現しないと、マウスは胚中心を作れなくなると同時にTFH細胞も存在しなくなります。
そこで、T細胞でBcl6が発現しないマウス(TFH細胞欠損マウス)と、B細胞でBcl6が発現しないマウス(胚中心欠損マウス)を作製しました。これら2種のマウスは、いずれもTFH細胞と胚中心の両方が存在しません。そして、これらのマウスと野生型マウスを使い、2009年の世界的大流行(パンデミック)の原因となった季節性インフルエンザウイルス(H1N1 Narita株)と、高病原性鳥インフルエンザウイルス(H5N1)に対する不活化ワクチン[9]の有効性を調べました。
実験では、それぞれのウイルスの致死量を①不活化ワクチン接種なしのTFH細胞マウスと胚中心欠損マウス、②不活化ワクチンを接種したTFH細胞欠損マウスと胚中心欠損マウス、③不活化ワクチンを接種した野生型マウスに感染させ、体重の変化を経時観察しました。その結果、不活化ワクチンを接種したTFH細胞欠損マウスと胚中心欠損マウスは、不活化ワクチンを接種した野生型マウスと同様にウイルスに対する強い抵抗性がみられ、極端な体重の変化はありませんでした(図1左)。また、不活化ワクチンを接種したTFH細胞欠損マウスの血中抗体価を測定したところ、IgG1抗体量は大きく減少しましたが、IgG2抗体量はわずかに減少しただけでした。(図1右)。
さらに、不活化ワクチンを接種したTFH細胞と胚中心の両方が存在しないマウス(TFH細胞欠損マウス)由来の血清[10](抗体が含まれる)を、ワクチンを接種したことのないTFH細胞欠損マウスに移入したところ、ウイルスに対する抵抗性を獲得しました。胚中心は、抗体にウイルスに対する高い親和性を与える免疫器官です。胚中心を持たないTFH細胞欠損マウスから得られた血清の親和性を試験管内で測定したところ、親和性は大きく低下していました。
これらの結果から、IgG2抗体はウイルスに対して親和性が低いにも関わらず、十分な中和活性を持っていると考えられます。つまり、ウイルスなどの抗原に対するワクチンの有効性は、抗体の親和性を上げなくても、低親和性のIgG2抗体を誘導することで十分に有効であることが分かりました。
次に共同研究グループは、インフルエンザウイルスに対して低親和性のIgG2抗体が、どのように産生されるのかを調べました。
まず、T細胞の関与を調べるために、T細胞を持たないマウスを作製しました。このマウスは、ウイルスに対して特異的なIgG抗体は全く産生されませんでした。
インフルエンザウイルスに対する免疫反応では、T細胞の一種であるⅠ型ヘルパーT細胞(TH1細胞)が活性化し、抗ウイルス性のサイトカイン(情報伝達物資)であるインターフェロン-ガンマ(IFN-γ)を産生することが知られています。そこで、IgG抗体の産生におけるTH1細胞の関与を調べるために、TH1細胞が産生するIFN-γが存在しないマウスを作製し、IgG抗体価を測定しました。すると、このマウスはワクチンを接種してもIgG2抗体が産生されませんでした。この結果から、ワクチンによって活性化するTH1細胞の重要性が明らかになりました。
次に、活性化したTH1細胞が、IFN-γ以外のサイトカインを産生するかどうかを調べた結果、抗体産生を助ける働きをする液性因子インターロイキン-21(IL-21)[11]を産生することが分かりました。そこで、IL-21を産生しないマウスを作製し、ウイルスに対するIgG2抗体価を測定したところ、IgG2抗体価が大きく減少していました。これらの結果から、IgG2抗体の産生過程には、TH1細胞が活性化し、IFN-γと共にIL-21を産生することが必要であることが明らかになりました。
さらに、TH1細胞がIgG2抗体の産生を助けるかを調べました。ワクチンを接種したマウスからTH1細胞だけを集め、ワクチンを接種していないマウスに移植しました。すると、TH1細胞を移植したマウスでは、IgG2抗体が大量に産生されました。また、このIgG2抗体をワクチンを接種していないマウスに移入したところ、マウスはウイルス抵抗性を獲得しました。
以上の結果から、ワクチンでTH1細胞を活性化することで産生されるIgG2抗体は、低親和性であるにも関わらず高親和性の抗体に引けを取らないほどの高い中和活性を持つことが証明されました。
今後の期待
本成果から、IFN-γとIL-21を産生するTH1細胞の助けによって、B細胞から産生されるIgG2抗体のウイルス感染防御における役割が明らかとなりました。IgG2抗体は、ウイルスに対する抵抗性が高いことが示されています。そのため、IgG2抗体が有意に産生される状況は、ウイルスに対する生体防御に有効であると考えられます。
胚中心依存的に起こる抗体産生は、親和性を上げるために時間を要し、高親和性の抗体ばかりが選択されてしまうため、抗体の特異性が限定される可能性があります。一方、TH1細胞は親和性が低い反面、広範なウイルスに対応できる抗体を敏速に生産できます。そのため、効率よくTH1細胞を活性化できるワクチンの開発は、毎年のように新しく出現するインフルエンザウイルスに対抗する上で新しいワクチン戦術として期待できます。
※共同研究グループ
理化学研究所 統合生命医科学研究センター
サイトカイン制御研究チーム
チームリーダー 久保 允人(くぼ まさと)(東京理科大学生命医科学研究所 教授)
研究員 宮内 浩典(みやうち こうすけ)
創薬抗体基盤ユニット
ユニットリーダー 竹森 利忠(たけもり としただ)
テクニカルスタッフ 石毛 晶子(いしげ あきこ)
研究員(研究当時) 加地 友弘(かじ ともひろ)
統合ゲノミクス研究グループ
グループリーダー 小原 収(おはら おさむ)
上級研究員 渡辺 貴志(わたなべ たかし)
上級研究員(研究当時) 土方 敦司(ひじかたあつし)
統合細胞システム研究チーム
チームリーダー 岡田 眞理子(おかだ まりこ)
研究員 井上 健太郎(いのうえ けんたろう)
東京理科大学 生命医科学研究所
研究員 原田 康代(はらだ やすよ)
国立感染症研究所
免疫部第四室
室長 高橋 宜聖(たかはし よしまさ)
研究員 安達 悠(あだち ゆう)
感染病理部
部長 長谷川 秀樹(はせがわ ひでき)
東京大学医科学研究所 ウイルス感染分野
教授 河岡 義裕(かわおかよしひろ)
特任准教授 福山 聡(ふくやま さとし)
博士課程 前村 忠(まえむら ただし)
原論文情報
- Kosuke Miyauchi, Akiko Sugimoto-Ishige, Yasuyo Harada, Yu Adachi, Yoshiko Usami, Tomohiro Kaji, Kentaro Inoue, Hideki Hasegawa, Takashi Watanabe, Atsushi Hijikata, Satoshi Fukuyama, Tadashi Maemura, Mariko Okada-Hatakeyama, Osamu Ohara, Yoshihiro Kawaoka, Yoshimasa Takahashi, Toshitada Takemori and Masato Kubo, "Protective neutralizing influenza antibody response in the absence of T follicular helper cells", Nature Immunology, doi: 10.1038/ni.3563
発表者
理化学研究所
統合生命医科学研究センター サイトカイン制御研究チーム
チームリーダー 久保 允人 (くぼ まさと)
(東京理科大学生命医科学研究所 教授)
研究員 宮内 浩典 (みやうち こうすけ)
 久保 允人
久保 允人
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
東京理科大学 研究戦略・産学連携センター
Tel: 03-5228-7440 / Fax: 03-5228-7441
ura [at] admin.tus.ac.jp(※[at]は@に置き換えてください。)
補足説明
- 1.胚中心
B細胞と濾胞樹状細胞からなる細胞が集合して免疫応答が起こり、組織内で抗体が作られると考えられる領域のことをB細胞濾胞という。胚中心はB細胞濾胞の中心部にできる球状の構造で、B細胞とT細胞が協調して活発に抗体が作られる。 - 2.リンパ濾胞型ヘルパーT細胞(TFH細胞)
ヘルパーT細胞の一種。主にインターロイキン-21(IL-21)とIL-4を産生し、胚中心の形成と免疫グロブリンG1(IgG1)抗体の産生に関わる。 - 3.免疫グロブリンG2抗体(IgG2抗体)
IgG抗体のサブクラスの一つで、生体内ではIgG1より強いウイルス防御活性を持つ。 - 4.インターフェロン-ガンマ(IFN-γ)
サイトカイン(情報伝達物資)の一種。主に、ヘルパーT細胞の一種であるTH1細胞によって産生される。 - 5.Ⅰ型ヘルパーT細胞(TH1細胞)
ヘルパーT細胞の一種。抗原の情報をB細胞へ伝えて抗体産生を誘導したり、免疫応答を誘導する液性因子を放出することにより、免疫反応の司令塔として働く。ウイルス抗原によって強力に誘導され、主に細胞性免疫に関わると考えられている。 - 6.T細胞
免疫細胞の一種。他の細胞と接触し、サイトカインと呼ばれる液性因子を分泌することで、B 細胞や他の免疫細胞の細胞分化や機能を調節する。分泌するサイトカインの種類や局在場所の違いによって機能が異なる。 - 7.B細胞
免疫細胞の一種。抗体(免疫グロブリン、lg)の産生を担う。lgがウイルスや細菌、毒素といった異物に特異的に結合して排除する。 - 8.2次リンパ組織
抗体産生や細胞性免疫反応に関与する末梢性リンパ組織で、リンパ節、脾臓、パイエル板、扁桃などがある。 - 9.不活化ワクチン
病原体となるウイルスなどの感染する能力を失わせたものを原材料として作られたワクチン。これに対して、病原体となるウイルスなどの毒性を弱めて病原性をなくしたものを原材料として作られたものを生ワクチンという。 - 10.血清
血液を常温で放置したとき、細胞成分と凝固成分が除かれてできる淡黄色透明の上澄み。免疫抗体や各種の栄養素、老廃物を含む。 - 11.インターロイキン21(IL-21)
サイトカインの一種。ヘルパーT細胞では主にTFH細胞によって産生されるが、一部のTH1細胞でも産生される。
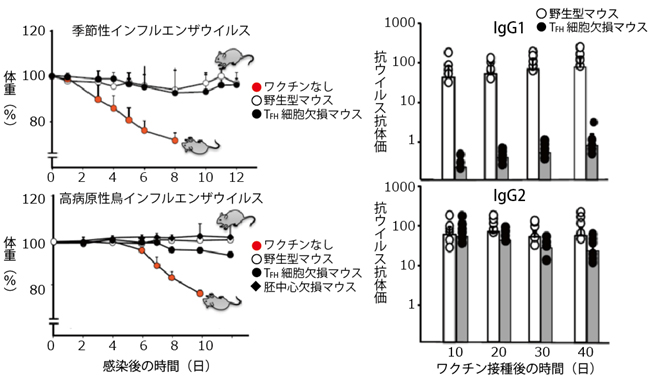
図1 マウスにおけるIgG2抗体によるウイルス感染防御
左上:季節性インフルエンザウイルスの致死量を感染させたマウスの体重変化。不活化ワクチン接種なしのTFH細胞欠損マウスの体重は減少したが、不活化ワクチンを接種した野生型マウスとTFH細胞欠損マウスの体重は、ほとんど減少しなかった。
左下:高病原性鳥インフルエンザウイルスの致死量を感染させたマウスの体重変化。上のグラフと同様に、不活化ワクチンを接種した野生型マウス、およびTFH細胞欠損マウスと胚中心欠損マウスの体重はほとんど減少しなかった。この結果はTFH細胞と胚中心の両方が存在しないTFH細胞欠損マウスと胚中心欠損マウスにおいても不活化ワクチンの有効性は損なわれなかったことを示している。
右:季節性インフルエンザウイルスに対する不活化ワクチン接種後に生成された抗ウイルス抗体価の時間変化。上のグラフはIgG1抗体価、下のグラフはIgG2抗体価。不活化ワクチンを接種した野生型マウスに比べてTFH細胞欠損マウスはIgG1抗体価が大きく減少したが、IgG2抗体価はわずかに減少しただけだった。この結果はTFH細胞と胚中心の両方が存在しないTFH細胞欠損マウスでも、不活化ワクチン接種によるIgG2抗体の産生は影響を受けないことを示している。
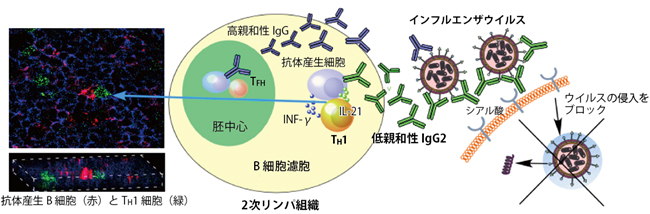
図2 IgG2抗体の産生機構とその役割
2次リンパ組織にあるB細胞濾胞において、ワクチンがTH1細胞(黄色)を活性化することで、インフルエンザウイルスに対して低親和性のIgG2抗体(緑色)の産生を誘導する。IgG2抗体はインフルエンザウイルスに結合して、体内への侵入過程を妨害する。TH1細胞は、2種類のサイトカインINF-γとIL-21を産生する。一方、胚中心ではB細胞とTFH細胞によって、高親和性のIgG抗体を誘導する。左の写真は、B細胞濾胞においてTH1細胞(緑色)が抗体産生B細胞(赤色)に接触して抗体産生を促すことを示している。