要旨
理化学研究所(理研)光量子工学研究領域超高速分子計測研究チームの倉持光研究員、田原分子分光研究室の竹内佐年専任研究員、田原太平主任研究員らの共同研究チーム※は、100兆分の1秒の光パルスを用いた独自の計測手法により、紅色光合成細菌[1]が持つタンパク質である青色光センサーが刺激に応答する瞬間の“最初の動き”を分子レベルで観測することに成功しました。
生物は外界からの光や熱などの刺激をタンパク質で構成されるさまざまなセンサーで検知し、それを生命活動の維持に役立てています。しかし巨大分子であるタンパク質において、その検知の仕組み、特に外的刺激に応答する瞬間の最初の動きを分子レベルで捉えるためには、分子の詳細な構造を区別し、かつ1兆分の1秒以内で起こる変化を追跡できる優れた観測手段が必要です。そのため、これまで最初の応答の解明は実験的に困難であると考えられてきました。しかし、初期過程は、その後の全ての生化学反応を運命づける誘因となる最も重要な過程です。そのため、高速の時間スケールでの分子の構造変化に関する情報が求められていました。
今回、共同研究チームは紅色光合成細菌の忌避走光性[2]という機能を担う青色光センサーである光受容タンパク質「イエロープロテイン(PYP)」に着目し、光照射直後の瞬間の分子構造とその後の変化を「フェムト秒時間分解インパルシブ・ラマン分光法」(フェムト秒は1,000兆分の1秒、10-15秒)と呼ばれる独自の最先端分光計測法を用いて調べました。この方法では、PYPに青色光を照射して変化を開始させた後に、わずか100兆分の1秒(10フェムト秒)の時間幅の光パルスで分子を瞬間的に揺さぶり、その揺れる様子から分子の形の変化の一部始終を“ストロボ写真”を撮るように観測しました。その結果、青色光の照射からわずか10兆分の1秒(0.1ピコ秒、1ピコ秒は1兆分の1秒)で光を吸収する部位の近くにある特定の水素結合が弱まり、それが歪んだ形状の分子を生じさせタンパク質の機能発現につながることを見いだしました。これはタンパク質が機能を発揮し始める精巧な仕組みをリアルタイムで追跡した画期的な成果といえます。
今後、この観測法を利用することにより、さまざまな光応答性タンパク質が機能する詳細な仕組みが明らかになるだけでなく、より優れた機能を持つ新しいタンパク質の設計・創製につながると期待できます。
本研究は、国際科学雑誌『Nature Chemistry』掲載に先立ち、オンライン版(2月6日付け:日本時間2月7日)に掲載されます。
※共同研究チーム
理化学研究所
光量子工学研究領域 エクストリームフォトニクス研究グループ
超高速分子計測研究チーム
研究員 倉持 光(くらもち ひかる)(理研 田原分子分光研究室 研究員)
田原分子分光研究室
専任研究員 竹内 佐年(たけうち さとし)
主任研究員 田原 太平(たはら たへい)
奈良先端科学技術大学院大学
教授 片岡 幹雄(かたおか みきお)
准教授 上久保 裕生(かみくぼ ひろなり)
大学院生 米澤 健人(よねざわ けんと)
背景
生物は外界からの光や熱などの刺激をさまざまなセンサーで検知し、それを生命活動の維持に役立てています。例えば、ヒトの視覚や植物の屈光性などもこうした検知に基づく生命活動です。センサーは、20種類のアミノ酸の鎖からなる巨大分子のタンパク質で構成されています。外部からの刺激に応答してタンパク質の構造は変化し、特定の構造をとることで初めてその機能が発揮されます。このように生物の機能と密接に関わっているタンパク質の構造を調べることは、その機能を理解するために本質的に必須であり、現代の化学・生物学において最も重要な課題の一つです。
タンパク質の中でも生物の光センサーとしての機能を担うのが、光受容タンパク質です。光受容タンパク質は、光の照射によってタンパク質の機能を容易に活性化することができます。そのため、機能が発現するまでのタンパク質の構造変化、動作機序を逐次的に調べることができる理想的な研究対象として、これまでさまざまな計測手法を用いて精力的に研究されてきました。多くの光受容タンパク質が機能を発現するまでの過程は、フェムト秒(1フェムト秒は1,000兆分の1秒、10-15秒)~数秒と非常に広い時間範囲にわたっています。特に、光が照射された直後のフェムト秒~ピコ秒(1ピコ秒は1兆分の1秒、10-12秒)の時間領域で起こる最初の構造変化を分子レベルで捉えることは、これまで実験的に困難でした。こうした初期過程は、その後の全ての生化学反応を運命づける誘因となる最も重要な過程です。そのため、高速の時間スケールでの分子の構造変化に関する情報が求められていました。
そこで、共同研究チームは紅色光合成細菌の忌避走光性という機能を担う青色光センサーである光受容タンパク質「イエロープロテイン(PYP)」を対象に、フェムト秒~ピコ秒にわたる機能発現初期過程における分子の構造変化を、100兆分の1秒(10フェムト秒)のパルス光を用いて解明することを試みました。
研究手法と成果
共同研究チームは、高速で変化する光受容タンパク質の構造を観測するために「フェムト秒時間分解インパルシブ・ラマン分光法」と呼ばれる独自に開発した分光計測法を用いました。まず、波長450ナノメートル(nm、1nmは10億分の1メートル)の青色のフェムト秒パルス光を用いて、PYPの光反応を開始させました。次に、生成された反応中間状態のPYPに100兆分の1秒(10フェムト秒)の時間幅を持った極短パルス光を照射し、インパルシブ誘導ラマン[3]と呼ばれる過程を利用して一斉に分子を揺さぶり、さらにこの分子の揺れる様子を時間的に遅れたもう一つの極短パルス光を使って実時間で観測しました(図1)。こうして観測される分子の振動には、分子を揺さぶった瞬間の構造に関する詳細な情報が含まれています。反応を開始させる青色光を照射してから分子を揺さぶる光を照射するまでのタイミングを変えながら測定を行うことで、これまで困難とされていたフェムト秒の時間領域におけるPYPの構造変化の追跡を行いました。
このようにしてPYPの反応初期過程における分子振動の変化を追跡したところ、まず光反応開始直後(0.1ピコ秒後、10兆分の1秒)に135cm-1の周波数を持つ分子の振動(約240フェムト秒周期)が明瞭に観測されました(図2A赤線)。ところがさらに0.15ピコ秒ずつ経過すると、この強い振動はほとんど観測されなくなることが分かりました(図2A緑線、青線)。フーリエ変換[4]による定量的な解析を行い、分子の揺れを振動スペクトル[5]の形にしてみると、135cm-1の振動に対応するバンド[6]が即座に減衰していく様子が分かりました(図2B)。そこで、変異体に対する参照実験との比較や計算化学から得られる知見と合わせて解析を行いました。その結果、観測された信号の減衰は、発色団[7]とそれに隣接するアミノ酸残基との間の水素結合強度の低下および水素結合構造の変化に由来することが分かりました。
また、光反応開始から1ピコ秒ほど経つと、最初にはみられなかった振動が徐々に現れてくる様子も観測されました。これはPYPの機能発現に必須である「I0状態」と呼ばれる光反応中間体に由来する振動であることが分かりました。I0状態が十分に生成する時間(反応開始後60ピコ秒後)まで待って分子を揺さぶったところ、これまで測定困難とされていたI0状態の振動スペクトルを高い信号対雑音比(S/N比)[8]で明瞭に得ることができました(図2C)。
反応開始前の発色団はトランス型構造をとっていますが(図1)、このI0状態の構造は最先端の時間分解X線結晶構造解析を用いた研究でも明確に決まらず、世界的な論争として大きな注目を集めてきました。今回、共同研究チームは量子化学計算などを駆使して測定されたスペクトルを解析し、トランス-シス異性化[9]を起こした直後のI0状態では発色団は、これまでには観測されたことのないようなねじれた分子構造をとっている、ということを分光学的に示しました。
今後の期待
今回、100兆分の1秒のパルス光を用いた独自の計測手法を用いることで、従来法では困難であった光を吸収した直後にタンパク質内で起こる非常に速い、小さな構造変化を観測することが可能になりました。本研究は、PYPの機能発現機構の全貌を分子レベルで解明するための大きな一歩になります。同時に、光受容タンパク質だけでなく、光によってさまざまな機能を発現する多くの物質において、極めて速く起こる最初の構造変化を検出することができること、またそれによって最初の小さな変化がその後の大きな構造変化や機能の発現をどのように引き起こすのかという問題に迫ることができる可能性を示しています。このような研究から得られる知見は、新しい光機能をデザインするための基盤を与えると期待できます。
また、本研究の対象である忌避走光性だけでなく、光合成、視覚、色認識、植物の屈光性など、生物学的に重要な多くの光反応の初期過程はフェムト秒~ピコ秒の速い時間領域で起こります。今後、本手法はさまざまな光応答性タンパク質が機能する詳しい仕組みを明らかにするだけでなく、より優れた機能を持つ新しいタンパク質の設計、創製につながると期待できます。
原論文情報
- Hikaru Kuramochi, Satoshi Takeuchi, Kento Yonezawa, Hironari Kamikubo, Mikio Kataoka, Tahei Tahara, "Probing the early stages of photoreception in photoactive yellow protein with ultrafast time-domain Raman spectroscopy", Nature Chemistry, doi: 10.1038/nchem.2717
発表者
理化学研究所
光量子工学研究領域 エクストリームフォトニクス研究グループ 超高速分子計測研究チーム
研究員 倉持 光(くらもち ひかる)
主任研究員研究室 田原分子分光研究室
専任研究員 竹内 佐年(たけうち さとし)
主任研究員 田原 太平(たはら たへい)
 倉持 光
倉持 光
 竹内 佐年
竹内 佐年
 田原 太平
田原 太平
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
お問い合わせフォーム
産業利用に関するお問い合わせ
理化学研究所 産業連携本部 連携推進部お問い合わせフォーム
補足説明
- 1.紅色光合成細菌
光エネルギーを利用して二酸化炭素を同化し生育する、つまり光合成を行う細菌を光合成細菌と呼ぶ。そのうち紅色光合成細菌は、電子供与体として硫化水素を利用するため、酸素は放出せず、またカロテノイドの蓄積により赤色や褐色を呈する。 - 2.忌避走光性
光が当たると、細菌が光の方向とは逆方向に自発的に動く機能のこと。青色の光や紫外光はエネルギーが高く、生物には有害なことが多い。そのため自己防御機能の一環として、ある種の菌はこれらの光を避ける機能を持つ。 - 3.インパルシブ誘導ラマン過程
100兆分の1秒(10フェムト秒)程度の非常に短い光パルスを分子に照射すると、分子内の原子が強制的に、一斉に振動を始める。これをインパルシブ誘導ラマン過程と呼ぶ。 - 4.フーリエ変換
さまざまな振動成分から構成されている信号から、各成分を抽出する数学的手法。時間領域の信号のフーリエ変換を行うと、元の信号にどのような周波数成分の振動がどのような割合で含まれているかという情報がスペクトルとして得られる。 - 5.振動スペクトル
多原子分子には各原子の動きの組み合わせにより、一定数の基準振動と呼ばれる分子の振動(伸び縮みの繰り返し運動)モードがある。振動スペクトルは分子がどのような振動数の基準振動を持つかを示す。これら基準振動の振動数はその分子の状態や周りの環境に対して鋭敏に変化するので、振動スペクトルからは分子構造に関する詳細な情報が得られる。 - 6.バンド
スペクトルの中に現れる信号強度が山形に増大した部分。 - 7.発色団
光受容タンパク質の中で実際に光を吸収する分子。PYPは125個のアミノ酸から構成されるタンパク質であるが、青色の光を吸収するのはタンパク質骨格に結合したトランス-パラクマル酸という発色団分子だけである。 - 8.信号対雑音比(S/N比)
一般的には測定時の信号(signal)と雑音(noise)の比率を示す。S/N比が大きいほど、精度の高い測定データが得られる。 - 9.トランス-シス異性化
分子式が同じで構造が異なる化合物を互いに構造異性体であるという。特に炭素・炭素原子間に二重結合を持つR1R2C=CR1R2の化合物などにおいて、同種の基(R1とR1、R2とR2)が二重結合について、同じ側にある配置をシス型、反対側にある配置をトランス型という。このトランス型とシス型が入れ替わることをトランス-シス異性化と呼ぶ。
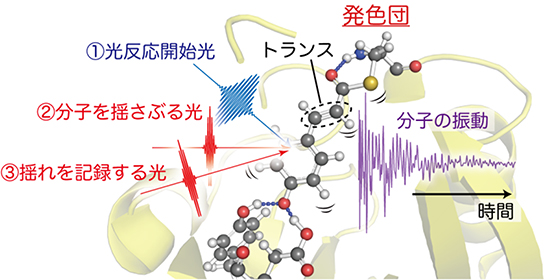
図1 イエロープロテインに対する「フェムト秒時間分解インパルシブ・ラマン分光法」概念図
紅色光合成細菌の青色光センサーである光受容タンパク質イエロープロテイン(PYP)に波長450nmの青色のフェムト秒パルス光を照射し、反応を開始させる(①)。次に反応中間状態のPYPに100兆分の1秒(10フェムト秒)の極短パルス光を照射することにより、分子を揺さぶる(②)。さらに、分子の揺れる様子を②と同様のもう1つの極短パルス光を使って実時間で観測する(③)。反応開始前の発色団には、トランス型の構造が存在する。
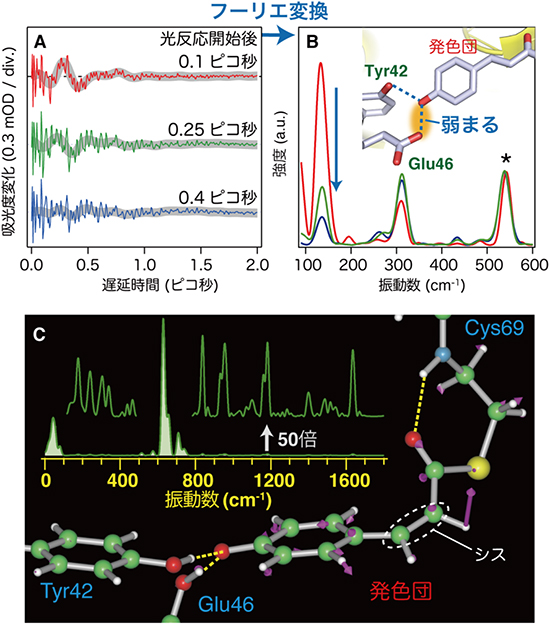
図2 PYPに対するフェムト秒時間分解インパルシブ・ラマン分光のデータ
A:PYPの反応過程における分子振動の変化。光反応開始後0.1ピコ秒では135cm-1(約240フェムト秒周期)の周波数の振動が明瞭に観測された(赤線)。しかし、さらに0.15ピコ秒ずつ経過すると、強い振動はほとんど観測されなくなった(緑線、青線)。灰色の太い線で表した波形が、135cm-1(約240フェムト秒周期)の振動成分に対応する。
B:Aの分子振動をフーリエ変換により振動スペクトルの形にすると、135cm-1の振動に対するバンドが即座に減衰していく様子が分かった。これは、青色の破線で示した、発色団とそれに隣接するアミノ酸残基との水素結合強度の低下および水素結合構造の変化に由来することも分かった。
C:上は、PYPの機能発現に必須なI0状態が十分に生成する時間(反応開始後60ピコ秒後)における振動スペクトル。0-100cm-1、600-700cm-1、700-750cm-1辺りに大きな振動バンドがみられる。また、100-500cm-1、750-1800cm-1の小さなバンドは50倍にしてその上に緑線で表している。下は、量子化学計算から示されるそのねじれた分子構造。反応開始前にトランス型であった発色団が異性化してねじれたシス型になっていることが分かる。