要旨
理化学研究所(理研)統合生命医科学研究センター骨関節疾患研究チームの池川志郎チームリーダー、小倉洋二客員研究員と側彎(そくわん)症臨床学術研究グループ[1]による共同研究グループ※は、思春期特発性側彎症(AIS:Adolescent Idiopathic Scoliosis)の進行、重症化に関連する新たな遺伝子「MIR4300HG」を発見しました。
側彎症は背骨が曲がる疾患です。神経や筋肉の病気、脊椎の奇形などが原因で起きるものもありますが、その多くは原因が特定できない特発性側彎症です。特発性側彎症の中で最も発症頻度が高いのが思春期に起きるAISで、全世界で人口の約2%にみられる非常に頻度の高い疾患です。AISの発症、進行には遺伝的要因が関与すると考えられ、世界中でその原因遺伝子の探索が行われてきました。先に池川チームリーダーらは、ゲノムワイド相関解析(GWAS)[2]を行い、AISの発症しやすさ(疾患感受性)を決定する遺伝子「LBX1」「GPR126」「BNC2」を発見しました注1-3)。しかし、AISの進行に関連する遺伝子はこれまで発見されていませんでした。
今回、共同研究グループは、日本人のAIS患者2,142人について、側彎の進行の有無を決定し、ヒトのゲノム全体を網羅する約750万個の一塩基多型(SNP)[3]を調べ、側彎の進行の有無と強い相関を示すSNPを見つけました。さらに、この結果を別の日本人のAIS患者606人を用いた相関解析により確認し、AISの進行と非常に強い相関を示す一群のSNPを見つけました。これらのSNPはMIR4300HGという遺伝子内に存在しました。MIR4300HGは、「MIR4300」というマイクロRNA[4]をコードしています。高い相関を示すSNPの一つrs35333564を含むゲノムの領域は、遺伝子の発現に関わるエンハンサー[5]活性を持ち、進行に関連するSNPの対立遺伝子座位(アレル)[6]を持つゲノムの領域では、エンハンサー活性が低下していることから、MIR4300量の低下が、側彎の進行に関連する可能性が考えられます。
今後、MIR4300の機能をさらに詳しく調べることで、側彎進行の予測法の開発、分子レベルでのAISの病態の解明、側彎進行の予防法や治療薬の開発が期待できます。
本成果は、『Human Molecular Genetics』掲載に先立ち、オンライン版(7月21日付)に掲載されました。
注1)2011年10月24日プレスリリース「思春期特発性側弯症の原因を解明、治療への大きな一歩」
注2)2013年5月13日プレスリリース「思春期特発性側彎症(AIS)発症に関連する遺伝子「GPR126」を発見」
注3)2015年7月24日プレスリリース「思春期特発性側彎症(AIS)発症に関連する遺伝子「BNC2」を発見」
※共同研究グループ
理化学研究所
統合生命医科学研究センター
骨関節疾患研究チーム
チームリーダー 池川 志郎(いけがわ しろう)
客員研究員 小倉 洋二(おぐら ようじ)
上級研究員 稲葉 郁代(いなば いくよ)
統計解析研究チーム
チームリーダー 鎌谷 洋一郎(かまたに よういちろう)
統合生命医科学研究センター
副センター長 久保 充明(くぼ みちあき)
背景
側彎(そくわん)とは、背骨が横に曲がった状態のことです(図1)。ヒトの背骨は完全に真っすぐではありませんが、曲がりの角度が10度以上になると病的(側彎症)と考えられています。曲がりの角度が20度を超えると、装具の着用など何らかの治療をする必要が生じ、40度を超えると、多くの場合手術治療が必要となります。さらに重度になった場合は、肺機能が低下し、腰痛や背部痛の発症が増加するとされています。進行すると治療が困難になるので、早期発見が大切です。
側彎症を引き起こす原因はさまざまで、神経麻痺や筋ジストロフィーなどの疾患により引き起こされることもありますが、多くは原因が特定できない特発性側彎症です。特発性側彎症は、発症時期などによりいくつかのタイプに分けられます。そのうち、最も発症頻度が高いのが、10歳以降に発症・進行する思春期特発性側彎症(AIS:Adolescent Idiopathic Scoliosis)で、全世界で人口の約2%にみられる発症頻度の非常に高い疾患です。日本人でも約2%にみられ、学校保健法により側彎の学校検診が義務付けられているほど、社会的かつ医療上、重大な問題となっています。
過去の疫学研究などから、AISは遺伝的因子と環境的因子の相互作用により発症する多因子遺伝病で、その発症、進行には遺伝的要因が深く関与することが明らかになっています。これまで世界中の研究グループがAISの原因遺伝子の探索を行ってきました。池川チームリーダーらは、2011年にAISのゲノムワイド相関解析(GWAS)を行い、AIS発症に関連する遺伝子「LBX1」を発見しました。さらに、2013年には二つめのAIS感受性遺伝子「GPR126」を、2015年には三つめのAIS感受性遺伝子「BNC2」を発見しています。
しかし、AISの進行や重症化に関連する遺伝子はこれまで発見されていませんでした。そこで、共同研究グループはAISの進行に関連する遺伝子の発見に挑みました。
研究手法と成果
共同研究グループは、2,142人の日本人のAIS患者集団の臨床データ、X線画像データを解析し、側彎が進行したグループ(進行群)と進行しなかったグループ(非進行群)とに分類しました。そして、この集団に対して、先に行ったヒトのゲノム全体を網羅する約50万個の一塩基多型(SNP)のジェノタイピングデータ[7]をもとに、imputation法[8]という遺伝統計学的手法を用いて、約750万個のSNPについて、進行群と非進行群との間での頻度の差を調べました。その結果、進行群で10カ所のゲノム領域に存在するSNPに強い相関を発見しました(図2A)。
これらのSNPについて、別の日本人AIS患者集団606人の遺伝子を用いて、進行群と非進行群との間で相関解析を行なったところ、11番染色体の長腕11q14.1上のゲノム領域で、AISの進行との非常に強い相関(P = 1.98×10-9)が確認されました。強い相関を示す一群のSNPは「MIR4300HG」という遺伝子のイントロン1[9]内に存在しました(図2B)。MIR4300HGは、「MIR4300」というマイクロRNAをコードしています。MIR4300HG遺伝子のイントロン6の一部から、MIR4300が産生されます(図2C)。マイクロRNAはさまざまな生命現象に関与するとされていますが、MIR4300の機能はよく分かっていません。これまでに側彎との関係を指摘されたこともありませんでした。
そこで、公開されているさまざまな転写制御に関するゲノムデータ、遺伝子発現データを用いて、MIR4300の機能に関連すると考えられるSNPの候補を絞り込み、それらの候補であるSNPの転写能を一つ一つ実験的に検討しました。その結果、AISの進行と高い相関を示すSNPの一つrs35333564を含むゲノムの領域にはエンハンサー活性があること、そして、rs35333564の進行に関連する対立遺伝子座位(アレル)を持つエンハンサーでは、活性が低下していることが分かりました。よって、MIR4300HG遺伝子の転写の低下によるMIR4300量の低下が、側彎の進行に関与する可能性が考えられます。今回の結果は、マイクロRNAと側彎症の関係を示した最初の例です。
今後の期待
AISの治療において、進行、重症化の予防は最重要課題ですが、これまであまり有効な方法がありませんでした。本研究におけるAIS進行関連遺伝子の発見は、より簡便、正確、かつ確実な側彎の進行の予測法の開発、分子レベルでのAISの病態の解明、画期的な側彎進行の予防法や治療薬の開発につながると期待できます。
原論文情報
- Yoji Ogura, Ikuyo Kou, Yohei Takahashi, Kazuki Takeda, Shohei Minami, Noriaki Kawakami, Koki Uno, Manabu Ito, Ikuho Yonezawa, Takashi Kaito, Haruhisa Yanagida, Kei Watanabe, Hiroshi Taneichi, Katsumi Harimaya, Yuki Taniguchi, Toshiaki Kotani, Taichi Tsuji, Teppei Suzuki, Hideki Sudo, Nobuyuki Fujita, Mitsuru Yagi, Kazuhiro Chiba, Michiaki Kubo, Yoichiro Kamatani, Masaya Nakamura, Morio Matsumoto, Japan Scoliosis Clinical Research Group, Kota Watanabe and Shiro Ikegawa., "Identification of MIR4300HG on chromosome 11q14.1 as a susceptibility gene for progression of adolescent idiopathic scoliosis ", Human Molecular Genetics, doi: 10.1093/hmg/ddx291
発表者
理化学研究所
統合生命医科学研究センター 骨関節疾患研究チーム
チームリーダー 池川 志郎(いけがわ しろう)
客員研究員 小倉 洋二(おぐら ようじ)
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
お問い合わせフォーム
産業利用に関するお問い合わせ
理化学研究所 産業連携本部 連携推進部
お問い合わせフォーム
補足説明
- 1.側彎症臨床学術研究グループ
慶應義塾大学医学部整形外科学教室渡邉航太専任講師を中心とする側彎症の専門医集団で構成された臨床学術研究グループ。主なメンバーは以下の通り。
慶應義塾大学医学部(松本守雄教授、中村雅也教授、渡邉航太専任講師、高橋洋平元助教、小倉洋二元助教(現理研客員研究員)、武田和樹助教、岡田英次朗助教、藤田順之専任講師、八木満専任講師)、北海道大学(伊東学特任教授(現国立病院機構北海道医療センター脊椎脊髄病センター長)、須藤英毅准教授)、聖隷佐倉市民病院(南昌平名誉院長、小谷俊明院長補佐、佐久間毅せぼねセンター長)、獨協医科大学(種市洋教授)、防衛医科大学(千葉一裕教授、細金直文講師)、獨協医科大学越谷病院(飯田尚裕准教授)、河野整形外科(河野克己院長)、順天堂大学(米澤郁穂准教授)、東京大学(谷口優樹助教)、聖マリアンナ医科大学(赤澤努教授)、新潟大学(渡辺慶講師)、金沢大学(出村諭講師)、名城病院(川上紀明院長補佐、辻太一医師)、奈良県立医科大学(重松英樹講師)、大阪大学(海渡貴司助教)、神戸医療センター(宇野耕吉副院長、鈴木哲平医師)、神戸大学(西田康太郎准教授、角谷賢一朗助教)、福岡市立こども病院(柳田晴久部長)、九州大学(播广谷勝三准教授) - 2.ゲノムワイド相関解析(GWAS)
疾患の感受性遺伝子を見つける方法の一つ。ヒトのゲノム全体を網羅する遺伝子多型を用いて、疾患を持つ群と疾患を持たない群とで遺伝子多型の頻度に差があるかどうかを統計学的に検定する方法。検定の結果得られたP値(偶然にそのようなことが起こる確率)が低いほど、相関が高いと判定できる。GWASは、Genome-Wide Association Studyの略。 - 3.一塩基多型(SNP)
ヒトゲノムは30億塩基対のDNAからなるが、個々人を比較するとそのうちの 0.1%の塩基配列の違いがある。これを遺伝子多型という。遺伝子多型のうち一つの塩基が、ほかの塩基に変わるものを一塩基多型(SNP:Single Nucleotide Polymorphismの略)と呼ぶ。 - 4.マイクロRNA
ゲノムから転写されるタンパク質をコードしていない21~25塩基程度の1本鎖RNA。標的となるタンパク質をコードしているmRNAの3’末端側の非翻訳領域に結合してその翻訳を阻害する。 - 5.エンハンサー
主に遺伝子の上流や下流に位置し、遺伝子の転写効率を変化させるゲノムの特定の配列のうち、プロモーターの転写効率を著しく高めるものをエンハンサー領域(配列)という。 - 6.対立遺伝子座位(アレル)
哺乳類は母親と父親から同じ遺伝子セットを持つ染色体を1組ずつ受け継ぐ。この両親から受け継いだ1対の遺伝子セットを対立遺伝子座またはアレルと呼ぶ。 - 7.ジェノタイピングデータ
実験的に決定された遺伝子多型の対立遺伝子座の配列データ。 - 8.imputation法
実験的に決定された遺伝子多型の頻度データをもとに、遺伝子多型間の連鎖不平衡を利用して、未知の遺伝子多型の頻度を推定する遺伝統計学的手法。 - 9.イントロン1
遺伝子の中でゲノムDNAからmRNAに転写される部分をエキソンと称する。ゲノムDNA上で、エキソンとエキソンの間のmRNAに転写されない部分をイントロンと称する。イントロン1は、一番先頭のエキソン(エキソン1)と次のエキソン(エキソン2)とに挟まれた領域。
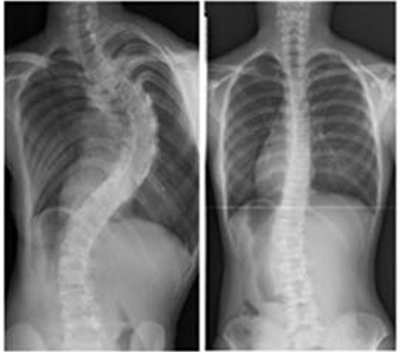
図1 側彎症の進行例と非進行例
左:進行例、右:非進行例。現在、進行を確実に予測する方法はない。
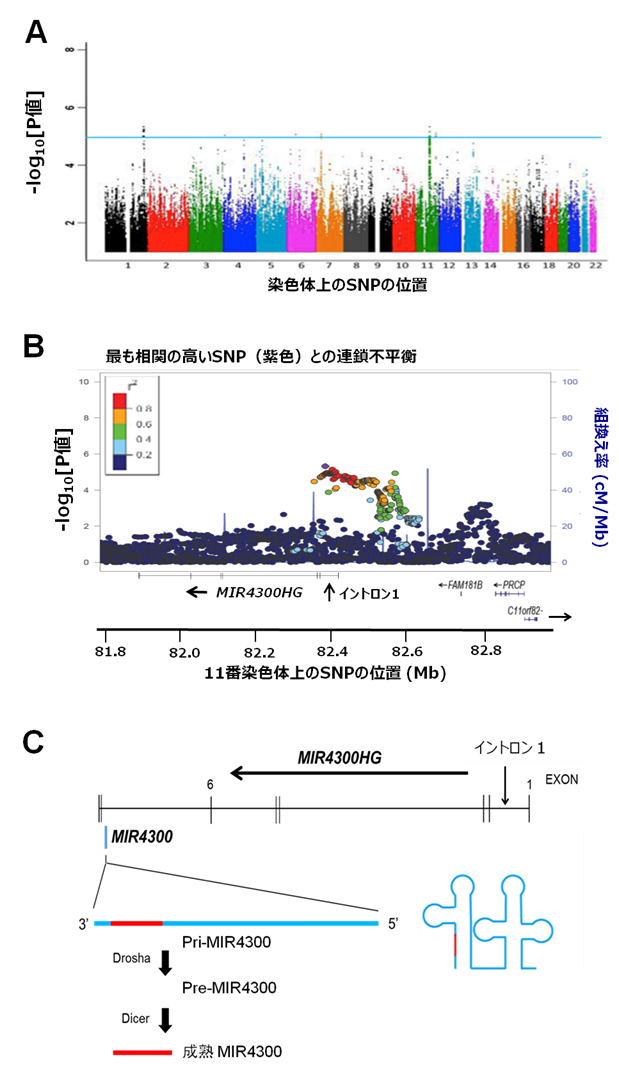
図2 相関解析の結果とMIR4300HG遺伝子、MIR4300の関係
A)2,142人の全ゲノム相関解析の結果。横軸はSNPの染色体ごとの位置。SNPを染色体ごとに色分けして示している。縦軸はP値の対数値で示した相関の強さ。青線(P=1.0x10-5)を超える強い相関を示すゲノム領域が10カ所存在する。
B)相関解析の結果の11番染色体長腕の部分(11q14.1)の拡大図。強い相関を示すSNPが、MIR4300HG遺伝子の上流からイントロン1にかけて存在する。
C)MIR4300HG遺伝子の遺伝子構造とMIR4300の位置、および成熟MIR4300(赤線の部分)が産生される過程。MIR4300HG遺伝子のイントロン6の一部からMIR4300ができる。その過程はまず、ヘアピン構造の長いRNA(Pri-MIR4300)として転写され、それをRNA分解酵素Droshaによって中間産物のPre-MIR4300が切り出される。Pre-MIR4300はさらに、別のRNA分解酵素Dicerによって切り出され、それが最終的に成熟MIR4300となり、機能するようになる。