要旨
理化学研究所(理研)環境資源科学研究センター機能開発研究グループのキム・ジュンシク基礎科学特別研究員と篠崎一雄グループディレクター、東京大学大学院農学生命科学研究科の篠崎和子教授らの共同研究チームは、植物の根の正常な伸長には「小胞体ストレス応答機構(UPR)[1]」に関わる二つの転写因子[2]が必要であることを明らかにしました。
地中に根を張る陸上植物は、自ら移動して最適な環境に移ることができません。そのため高温・乾燥・病害など、さまざまな環境ストレスに対処するための複雑なストレス応答機構[3]を発達させてきました。植物は環境ストレスによって生長全般が抑えられます。しかし、特定の組織に対する生長制御に関してはよく分かっていませんでした。例えば、ストレス応答機構の一つであるUPRについて、モデル植物のシロイヌナズナでは、三つのbZIP型転写因子がUPRを制御していること、このうちbZIP28とbZIP60は制御のメジャー因子としてさまざまな環境ストレスに対する耐性付与していることは分かっています。しかし、残る一つのbZIP17の機能はよく分かっていませんでした。
今回、共同研究チームは、シロイヌナズナを用いてbZIP17を含む多重欠損変異株の作出を試みました。その結果、転写因子bZIP17とbZIP28の二重欠損変異株の作出に成功しました。各bZIPの単一欠損変異株が野生株と変わらない発生・生長を示すのに対して、この二重欠損変異株では、根が野生株のわずか10%程度までしか伸長せず、また複数の細胞伸長遺伝子の発現が抑制されていました。これらの結果は、植物のUPRが本来の機能である環境ストレス応答だけでなく、根の伸長に対する特別な制御機能を持つことを示しています。
今後、UPRによる根の伸長制御の分子機構をさらに詳しく解析することで、小胞体ストレスシグナルを含めたメカニズムの理解が深まると考えられます。また、bZIPと根の伸長を結びつける原因遺伝子を究明することで、作物の改良に役立つ技術開発につながると期待できます。
本研究成果は、国際科学雑誌『Plant Physiology』オンライン版(1月24日付け:日本時間1月25日)に掲載されました。
背景
地中に根を張る陸上植物は、自ら移動して最適な環境に移ることができません。そのため、高温、乾燥、病害などさまざまな環境ストレスに対処するための複雑なストレス応答機構を発達させてきました。植物は環境ストレスによって、生長全般が抑えられます。しかし、特定の組織に対する生長制御に関してはよく分かっていませんでした。
小胞体は、真核生物の細胞内に存在する細胞小器官[4]の一種で、生命現象の維持に必須なタンパク質の加工(折り畳み)や糖鎖などの修飾が行われ、細胞の工場に例えられます。タンパク質の加工プロセスは外部環境の変化にとても敏感で、高温、乾燥、病害などのさまざまな刺激を受けると、正しく加工されていない変性タンパク質を発生させます。小胞体は過剰な変性タンパク質の蓄積をストレスの一種と認知し、その程度に応じてタンパク質加工プロセスを強化する遺伝子発現制御経路を持っています。「小胞体ストレス応答機構(UPR)」と呼ばれるこの機構は、さまざまな種類の環境ストレスに応答するため、細胞が外部環境を統合的に判断するバロメーターの一つとして機能していると考えられます。
代表的なモデル植物であるシロイヌナズナでは、三つのbZIP型転写因子(bZIP17、bZIP28、bZIP60)がUPRを制御しています。bZIP28とbZIP60は、UPR制御のメジャー因子として、植物にさまざまな環境ストレスに対する耐性を付与することが明らかになっています。しかし、bZIP17の機能についてはよく分かっていませんでした。
研究手法と成果
共同研究チームは、bZIP17のUPR制御における機能を明らかにするために、モデル植物のシロイヌナズナを用いて、bZIP17を含む多重欠損変異株の作出を試みました。その過程において、これまで致死として報告されていたbZIP17とbZIP28の二重欠損変異株(bz17/28)の作出に成功しました。その結果、各bZIPの単一欠損変異株が野生株(WT)と変わらない発生・生長を示すのに対して、bz17/28変異株は野生株に比べて生育速度が遅く矮性(背丈が低い)を示しました(図1A)。特に、根部の伸長における表現型の異常が著しく、野生株に比べてわずか10%程度までしか伸長しませんでした(図1B-D)。
転写因子は下流の遺伝子発現を制御するため、二つの転写因子をなくしたbz17/28変異株では、根の伸長に重要な未知の遺伝子の発現が欠如していると予測されました。その未知の遺伝子を探すため、bz17/28変異株と野生株間の網羅的遺伝子発現解析[5]を行い、両者の間で発現の差がある遺伝子を調べました。その結果、複数の細胞伸長遺伝子が、bz17/28変異株において発現抑制されていることが分かりました。既知のUPRの下流遺伝子は、小胞体ストレスに敏感に応答して発現が上昇しますが、これらの細胞伸長遺伝子はそのような応答性を示しませんでした。
この結果は、植物がbZIP17とbZIP28を介して既知のUPR制御とは異なる経路で複数の細胞伸長遺伝子の発現を維持し、その結果、根の正常な伸長を維持していることを示しています(図2)。
今後の期待
本成果は、植物の根の伸長制御にUPRが関与することを示し、根の伸長に関する新たな制御メカニズムを提案します。特に、UPRが外部環境を統合的に感知する機構であることを踏まえると、提案した制御メカニズムは野外の激しい環境にさらされる植物が示す最適な生長戦略を理解するための重要なモデルといえます。
今後、UPRによる根の伸長制御の分子機構をさらに詳しく解析することで、小胞体ストレスシグナルを含めたメカニズムの理解が深まると考えられます。また、現在計画しているbZIPと根の伸長を結びつける原因遺伝子の究明は、その学問的価値に止まらず、根を主に利用する作物の改良に役立つ技術開発にも寄与すると期待できます。
原論文情報
- June-Sik Kim, Kazuko Yamaguchi-Shinozaki, and Kazuo Shinozaki, "ER-Anchored Transcription Factors bZIP17 and bZIP28 Regulate Root Elongation", Plant Physiology, doi: 10.1104/pp.17.01414
発表者
理化学研究所
環境資源科学研究センター 機能開発研究グループ
基礎科学特別研究員 キム ・ジュンシク (Kim・June-Sik)
グループディレクター 篠崎 一雄 (しのざき かずお)
東京大学大学院 農学生命科学研究科 応用生命化学専攻
教授 篠崎 和子 (しのざき かずこ)
 キム・ジュンシク
キム・ジュンシク
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
お問い合わせフォーム
東京大学大学院農学生命科学研究科・農学部
総務課総務チーム(広報情報担当)
Tel: 03-5841-5484 / Fax: 03-5841-8196
koho [at] ofc.a.u-tokyo.ac.jp(※[at]は@に置き換えてください。)
産業利用に関するお問い合わせ
理化学研究所 産業連携本部 連携推進部
お問い合わせフォーム
補足説明
- 1.小胞体ストレス応答機構(UPR)
小胞体が過剰な変性タンパク質の蓄積をストレスの一種と認知し、その程度に応じてタンパク質加工プロセスを強化させる遺伝子発現制御経路のこと。UPRはunfolded protein responseの略。 - 2.転写因子
一つの細胞に存在する数万の遺伝子は、適材適所で機能するためにその発現が制御されねばならない。転写因子はタンパク質の一種で、DNAと直接相互作用することで標的遺伝子の発現を抑制、または誘導する。各転写因子は固有のターゲット塩基配列を持っていて、細胞内外の状況に応じて適切な遺伝子の発現制御に寄与する。 - 3.ストレス応答機構
生物の成長に不適切な環境、または病害などの被害から身を守るため、生物はさまざまなストレスに適切に対応するため複雑なストレス応答機構を発達させてきた。ストレス応答機構は、ストレスの受容・シグナルの伝達・対応遺伝子の活性化の三つの段階で構成されている。 - 4.細胞小器官
オルガネラともいう。細胞内の膜に囲まれた構造体で、それぞれが定められた機能を果たす。具体的には細胞核、小胞体、ゴルジ体、エンドソーム、リソソーム、ペルオキシソーム、ミトコンドリア、葉緑体、液胞などがある。 - 5.網羅的遺伝子発現解析
一つの細胞には数万から数十万の遺伝子が存在する。これらの全遺伝子の発現を網羅的に解析するためのさまざまな方法が開発されている。近年では比較的短い(100~300塩基)配列断片を大量(100億塩基以上)に生産する次世代シーケンス技術の利用が主流になっている。

図1 シロイヌナズナの二重欠損変異株(bz17/28)が示す表現型
- (A)bz17/28は、野生株(WT)に比べて背丈が低い矮性を示した。
- (B)単一欠損変異体のbz17、bz28は野生株と変わらないが、bz17/28は根部が著しく伸長しなかった。
- (C–D)bz17/28から見いだされた葉の異常発生および根部の拡大図。
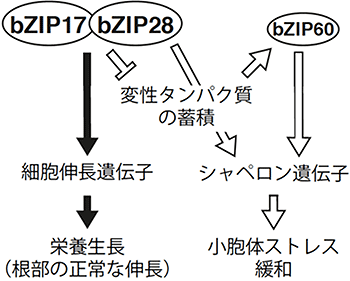
図2 本研究から提案された植物の小胞体ストレス応答機構(UPR)制御モデル
転写因子bZIP28とbZIP60はこれまでの知見通りのUPR、変性タンパク質の蓄積に応じて、タンパク質の正しい折り畳みを助けるシャペロン遺伝子を活性化する経路を制御する(白色の矢印)。一方、bZIP17はbZIP28と共同で、別の制御機構を通して細胞伸長遺伝子の発現を活性化することで、植物の栄養生長に寄与する(黒色の矢印)。