2018年3月8日
理化学研究所
農業・食品産業技術総合研究機構
極限的な乾燥耐性をつかさどる制御因子を同定
-カラカラに干からびても死なない昆虫の秘密の一端が明らかに-
要旨
理化学研究所(理研)予防医療・診断技術開発プログラムのオレグ・グセフマネージャー、農業・食品産業技術総合研究機構(農研機構)生物機能利用研究部門の黄川田隆洋上級研究員、リシャー・コルネット主任研究員らの国際共同研究グループ※は、ネムリユスリカ[1]という干からびても死なない昆虫を用いて、熱ショック転写因子[2]であるHSF1が極限的な乾燥耐性を制御することを明らかにしました。
アフリカ中部の半乾燥地帯に生息するネムリユスリカは「乾燥無代謝休眠[3]」と呼ばれる珍しい能力を持つ昆虫で、体内の水分のほとんどがなくなるほど干からびても死なないことで知られています。これまで、ネムリユスリカの乾燥耐性機構を理解するために、乾燥耐性関連因子の同定やゲノム構造の解読が進められてきました。しかし、乾燥耐性機構を制御する遺伝子発現ネットワークについてはほとんど分かっていませんでした。
今回、国際共同研究グループは、ネムリユスリカとその近縁で乾燥耐性がないヤモンユスリカのゲノムの比較から、ネムリユスリカでは乾燥で発現が上昇する遺伝子の転写開始点近傍に、ゲノム特異的なDNAモチーフ(TCTAGAA)が多く、かつ偏って存在すること、プロモーター領域[4]にTCTAGAAを持つ乾燥誘導性遺伝子には乾燥耐性関連遺伝子が多く含まれることを見いだしました。また、TCTAGAAはHSF1の結合領域に酷似していました。そこで、ネムリユスリカの培養細胞(Pv11細胞[5])を用いて、Hsf1遺伝子を機能抑制したところ、乾燥耐性関連遺伝子の発現が減少しました。さらに、Hsf1の発現を抑制したPv11細胞を乾燥させると、通常のPv11細胞を乾燥させたものと比べて、再び水に浸けた後の生存率が低下しました。すなわち、HSF1が乾燥耐性を制御する重要な転写因子であることが明らかとなりました。これらのことから、ネムリユスリカは進化の過程で、熱ストレス応答性の遺伝子発現制御ネットワークを転用化[6]した結果、極限的な乾燥耐性を発揮するようになった可能性が示されました。
今後、ネムリユスリカの乾燥無代謝休眠に関連した遺伝子群を任意の細胞で発現させることで、細胞や組織の常温乾燥保存法の開発が進みます。また、新しい乾燥耐性因子の同定にもつながると期待できます。
本研究は、米国の科学雑誌『Proceedings of the National Academy of Sciences of the United States of America』に掲載されるのに先立ち、オンライン版(2月20日付け:日本時間2月21日)に掲載されました。
本研究は、日本学術振興会(JSPS)科学研究費補助金 若手研究(A)「Mechanisms of anhydrobiosis-associated gene regulation in the cells and tissues of chironomid Polypedilum vanderplanki.(研究代表者:オレグ・グセフ)」、基盤研究(C)「ネムリユスリカの極限乾燥耐性におけるトレハロースの複合的な役割の解明(研究代表者:リシャー・コルネット)」、挑戦的萌芽研究「乾燥・塩ストレス制御型タンパク質発現システムの開発(研究代表者:黄川田隆洋)」および農林水産省農林水産技術会議−ロシア科学基金(MAFF/AFFRC-RSF)国際共同研究パイロット事業などの支援を受けて行われました。
※国際共同研究グループ
理化学研究所 予防医療・診断技術開発プログラム
マネージャー オレグ・グセフ(Oleg Gusev)
農業・食品産業技術総合研究機構 生物機能利用研究部門
上級研究員 黄川田 隆洋 (きかわだ たかひろ)(東京大学大学院 新領域科学研究科先端生命専攻 客員准教授)
主任研究員 リシャー・コルネット(Richard Cornette)
JSPS特別研究員(研究当時)十亀 陽一郎(そがめ よういちろう)(現 福島工業高等専門学校 特命助教)
カザン大学
主任研究員 エレナ・シャギマルダノワ(Elena Shagimardanova)
研究員 オルガ・コズロワ(Olga Kozlova)
研究員 アレクサンドル・チェルカソフ(Alexander Cherkasov)
東京大学大学院 新領域科学研究科 先端生命専攻
大学院生 徳本 翔子 (とくもと しょうこ)
東京工業大学 バイオ研究基盤支援総合センター
博士研究員 宮田 佑吾 (みやた ゆうご)
スコルコボ科学技術大学
教授 ミハイル・ゲルファンド(Mikhail S. Gelfand)
研究員 パベル・マジン(Pavel V. Mazin)
大学院生 ビタ・ステパノワ(Vita V. Stepanova)
モスクワ大学
研究員 マリア・ロガチェワ(Maria Logacheva)
研究員 アレクセイ・ペニン(Aleksey Penin)
ジョンホプキンス大学
博士研究員 アレクセイ・スツプニコフ(Alexey Stupnikov)
ローレンス・バークレー国立研究所
ソフトウェア開発者 ロマン・ストルミン(Roman Sutormin)
背景
全ての生物にとって、水は生命の基盤です。このため生物は、乾燥に対抗する適切な生命戦略を進化させてきました。アフリカ中部の半乾燥地帯に生息するネムリユスリカという昆虫は、体内の水分のほとんどがなくなるほど干からびても死なないことで知られています。この「乾燥無代謝休眠」と呼ばれる能力は、生命科学分野で一般的なモデル生物であるショウジョウバエやマウスなどにはありません。昆虫ではネムリユスリカの幼虫で確認されている能力です。これまで、ネムリユスリカの極限的な乾燥耐性機構を理解するために、乾燥耐性をもたらすタンパク質の同定やゲノム構造の解読が進められてきました。しかし、乾燥耐性機構を制御する遺伝子発現ネットワークについてはほとんど分かっていませんでした。
そこで、国際共同研究グループは、ネムリユスリカと近縁で乾燥耐性がないヤモンユスリカのゲノムを比較することで、ネムリユスリカに極限的な乾燥耐性機能の発現を制御する分子機構の解明を試みました。
研究手法と成果
国際共同研究グループはまず、ネムリユスリカとヤモンユスリカの解読済みのドラフトゲノムを比較しました。その結果、ヤモンユスリカに比べてネムリユスリカでは、乾燥によって発現上昇する遺伝子の転写開始点近傍に、ゲノム特異的なDNAモチーフ(TCTAGAA)が多く、かつ偏って存在することを見いだしました(図1)。プロモーター領域にこのモチーフを持つ乾燥誘導性の遺伝子には、LEA遺伝子やチオレドキシン遺伝子、トレハロース合成酵素遺伝子など、ネムリユスリカの乾燥耐性に関連した遺伝子が多く含まれていました。また、このモチーフは、ショウジョウバエの熱ショック転写因子(HSF1)の結合領域に酷似していました。
そこで、ネムリユスリカの培養細胞(Pv11細胞)を用いて、RNA干渉(RNAi)法[7]によってHsf1遺伝子を機能抑制したところ、上記の乾燥耐性関連遺伝子の発現が減少しました。さらに、Hsf1の発現を抑制させたPv11細胞は乾燥させると、通常のPv11細胞を乾燥させたものと比べて、有意に再水和後(再び水に浸けた後)の生存率が低下しました(図2)。このことから、HSF1がネムリユスリカの乾燥耐性を制御する重要な転写因子であることが明らかとなりました。
HSF1は、生物界に広く共通して存在する熱ストレス応答制御因子です。乾燥耐性がないヤモンユスリカもHSF1を持っていますが、ネムリユスリカのように乾燥耐性関連遺伝子の発現を誘導する機能はありません。したがって、進化の過程で、熱ストレス応答性の遺伝子発現制御ネットワークを転用化(co-option)した結果、ネムリユスリカは極限的な乾燥耐性を発揮できるようになった可能性が示されました。
今後の期待
本研究では、熱ショック転写因子HSF1がネムリユスリカの乾燥耐性を制御する重要な転写因子であることを明らかにしました。この成果を発展させると、さまざまな実用的な成果へつながる可能性があります。例えば、ネムリユスリカの乾燥無代謝休眠に関連する遺伝子群を任意の細胞で発現させることで細胞や組織の常温乾燥保存法の開発が加速します。
乾燥過程でHSF1によって発現が制御されている遺伝子は、極限的な乾燥耐性に寄与している遺伝子の可能性が高いといえます。すなわち、HSF1をノックアウトしたネムリユスリカ細胞で発現が大きく低下する遺伝子を探索することで、新しい乾燥耐性因子を同定できる可能性があります。
また、現在は遺伝子を細胞に発現させる際、薬剤処理や温度刺激を与える方法が主流となっています。今後、ネムリユスリカのHsf1遺伝子発現制御機構を応用することにより、乾燥刺激で任意の細胞に自由自在に遺伝子を発現できるシステムを構築できる可能性があります。
原論文情報
- Pavel V. Mazin, Elena Shagimardanova, Olga Kozlova, Alexander Cherkasov, Roman Sutormin, Vita V. Stepanova, Aleksey Stupnikov, Maria Logacheva, Aleksey Penin, Yoichiro Sogame, Richard Cornette, Shoko Tokumoto, Yugo Miyata, Takahiro Kikawada, Mikhail S. Gelfand, and Oleg Gusev, "Cooption of heat shock regulatory system for anhydrobiosis in the sleeping chironomid Polypedilum vanderplanki", Proceedings of the National Academy of Sciences of the United States of America (PNAS), doi: 10.1073/pnas.1719493115
発表者
理化学研究所
予防医療・診断技術開発プログラム
マネージャー オレグ・グセフ (Oleg Gusev)
農業・食品産業技術総合研究機構 生物機能利用研究部門
上級研究員 黄川田 隆洋 (きかわだ たかひろ)
主任研究員 リシャー・コルネット (Richard Cornette)
 オレグ・グセフ
オレグ・グセフ
 黄川田 隆洋
黄川田 隆洋
 リシャー・コルネット
リシャー・コルネット
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
お問い合わせフォーム
産業利用に関するお問い合わせ
理化学研究所 産業連携本部 連携推進部
お問い合わせフォーム
補足説明
- 1.ネムリユスリカ
干からびても死ぬことなく、水をかけると成長を再開する能力を持つ昆虫。この乾燥耐性は幼虫期のみに認められる。ナイジェリアなどのアフリカ北部の半乾燥地帯(サヘル地帯)の水たまりに生息する。学名は、 Polypedilum vanderplanki。最近、アフリカ南部(マラウィ)の半乾燥地帯に生息する同属のマンダラネムリユスリカ( Polypedilum pembai)も同じ能力を持つことが明らかになった。 - 2.熱ショック転写因子
転写因子の一つ。元々は、熱ショックタンパク質(HSP)をコードする遺伝子の発現を調節するものとして同定された。細胞に温度ストレスが加わると活性化し、ゲノム上の特定の配列に結合する。すると、その結合部位の近傍にある遺伝子の発現が上昇する。 - 3.乾燥無代謝休眠
干からびても死なない能力。英語では、anhydrobiosis(“水がない状態の生命”の意味)と呼ばれる。ネムリユスリカ以外では、細菌、酵母、カビの胞子、線虫、ワムシ、クマムシ、アルテミアの耐久卵、植物の種子、復活草などに認められる。脊椎動物には、この能力を持つ生物は存在しない。 - 4.プロモーター領域
遺伝子を発現させる機能を持つ塩基配列。プロモーターがないと遺伝子は発現しない。 - 5.Pv11細胞
ネムリユスリカの胚子由来の培養細胞。600mMという高濃度のトレハロースに浸漬させると、細胞の乾燥耐性関連遺伝子が発現し、常温乾燥可能な状態になる。乾燥させたPv11細胞は、冷蔵することなく1年程度の常温保存が可能である。なお、同様なトレハロース処理を行っても、Pv11以外の細胞は乾燥耐性能力の向上は認められない。このことから、ネムリユスリカ特有の遺伝子発現制御系が、乾燥耐性機能を発揮させるのに重要なことが示唆されていた。 - 6.転用化
進化の過程で、元々あった機能に加えて、ある遺伝子が別の機能を持つように変化することを指す。英語では、co-optionと呼ばれる。端的に言えば、「遺伝子の使い回し」を意味する。 - 7.RNA干渉(RNAi)法
特定の配列を持つRNAを細胞へ導入することで、相補的なmRNAの分解を促進させること。結果的に、その目的の遺伝子の機能を抑制することができる。
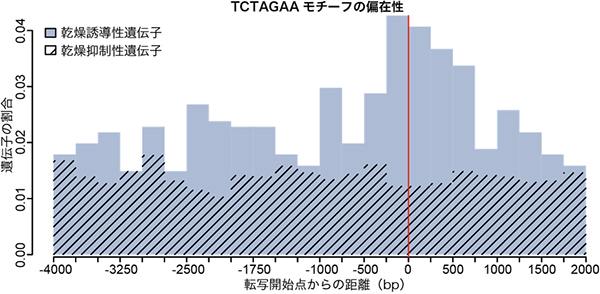
図1 ネムリユスリカの乾燥誘導性遺伝子の転写開始点近傍の特異的なDNAモチーフ
熱ショック転写因子HSF1の結合モチーフ様配列(TCTAGAA)は、ネムリユスリカの乾燥誘導性遺伝子の転写開始点(赤線)の近傍(±1,000 塩基以内)に偏って存在する。一方、乾燥抑制性遺伝子の転写開始点近傍ではモチーフの存在に偏りがない。
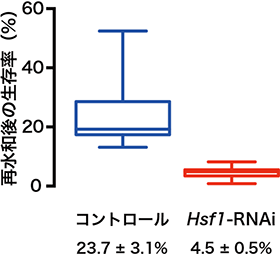
図2 Hsf1遺伝子を発現抑制したPv11細胞を乾燥させ再水和した後の生存率
RNA干渉(RNAi)法でHsf1遺伝子を発現抑制させたネムリユスリカ培養細胞(Pv11)は、乾燥耐性能力が減少する。Hsf1のsiRNAをエレクトロポレーションで導入したPv11細胞を10日間常温乾燥させ再水和した後の細胞生存率は、対照のPv11細胞と比べて約1/5に低下した。