2010年12月9日
独立行政法人 理化学研究所
独立行政法人 科学技術振興機構
細胞のストレス応答機構の分子メカニズムが明らかに
-ストレスによるIP3受容体の機能破壊が、神経細胞死による脳障害を引き起こす-
ポイント
- タンパク質GRP78がIP3受容体を介した細胞内カルシウム情報伝達系を制御
- ストレス条件下で、1型IP3受容体のカルシウム放出機能が低下し、細胞死を誘導
- 細胞死を誘導するという定説を覆し、IP3受容体がストレスから脳を守る
要旨
独立行政法人理化学研究所(野依良治理事長)と独立行政法人科学技術振興機構(以下JST、北澤宏一理事長)は、細胞内カルシウム濃度を調節するタンパク質IP3受容体(IP3R)が小胞体ストレスによって破壊され、神経細胞死※1を誘導することを世界で初めて発見しました。この発見は、IP3Rが細胞死を誘導するという従来の定説を覆すもので、ストレスによる神経変性疾患の発症メカニズムの理解につながります。これは、理研脳科学総合研究センター(利根川進センター長)発生神経生物研究チームの御子柴克彦チームリーダー(独立行政法人科学技術振興機構 戦略的創造研究推進事業 発展研究(SORST)「カルシウム振動プロジェクト」研究総括)、肥後剛康研究員(現、認知機能表現研究チーム)らによる成果です。
現代は、ストレス社会といわれて久しく、職場、家庭、学校での過度なストレスによる体調不良、心の病、自殺が社会的に大きな問題となっています。健康的な生活を過ごす上で、いかにストレスを軽減し、上手に付き合うかが、多くの人々にとって切実な問題となっています。細胞も同様に、絶えずストレスにさらされており、過度なストレスは、細胞が生来備えているストレス応答機構を破綻させ、細胞自らが死を選択する細胞死(アポトーシス)を引き起こすことが分かっています。特に、神経細胞がストレスにさらされると、細胞死による脳機能の低下、ひいてはさまざまな神経変性疾患を引き起こすと考えられています。ストレスによって神経細胞のストレス応答機構がどのように破綻し、細胞死が誘導されるかを解明することができると、神経変性疾患などの治療法の確立に大いに貢献するにもかかわらず、いまだにストレス応答機構破綻の分子メカニズムは明らかとなっていません。今回、研究チームは、細胞内のカルシウム濃度を調節するタンパク質であるIP3Rの機能が小胞体ストレスによって破壊され、神経細胞死を誘導することを世界で初めて発見しました。この発見は、ある特定のタンパク質(IP3R)がストレスから脳を守る働きをしていることを示唆したという点で画期的なものであるとともに、IP3Rが細胞死を誘導するという定説を覆すこととなりました。今後は、学術的貢献だけにとどまらず、ストレスによる神経変性疾患の発症メカニズムの理解や神経変性疾患の発症予防を含む治療への応用に貢献すると期待されます。
本研究成果は、米国科学雑誌『Neuron』(ニューロン)(12月9日号)に掲載され、その表紙を飾ります。
背景
細胞は、絶えず酸化ストレスや虚血などの外的なストレスにさらされているだけでなく、小胞体ストレスなどの細胞内のストレスにもさらされています。小胞体ストレスとは、異常なタンパク質が、その合成の場である細胞内小器官(小胞体)の内部に蓄積してしまう状態のことで、単細胞生物から高等ほ乳動物まで存在する現象です。多くの生物種において、進化的に小胞体ストレスへの応答機構が発達していますが、一方で、強度の小胞体ストレスは、それら応答機構を破綻させ、最終的には細胞死を誘導することが分かっています。また、ヒトでは、小胞体ストレスによる細胞死が、神経変性疾患や糖尿病などさまざまな疾患に関与すると考えられています。しかし、その発症のメカニズムはほとんど明らかとなっていません。
生物の細胞は、細胞外からの刺激が適切で許容範囲内であれば、細胞応答に必要な情報に変換することができます。変換される情報の代表的なものとして、細胞内のカルシウムが挙げられます。カルシウムは、細胞内でイオンとして存在するため、迅速かつ広範囲にわたる濃度変化、つまり空間移動が可能です。その結果、多彩な情報伝達が生み出され、細胞分裂、細胞死、受精、発生などのさまざまな細胞応答の制御が可能となります。通常、細胞内のカルシウム濃度は極めて低く抑えられており、細胞内カルシウム濃度上昇とその変動は、小胞体からのカルシウムの放出に依存しています。細胞質へのカルシウム放出は、小胞体に局在するカルシウムを放出するタンパク質であるIP3受容体(IP3R)を介して行われています。IP3Rは、ホルモンや神経伝達物質などの細胞外刺激で細胞内に産生されるIP3が結合することで活性化され、カルシウムを小胞体内腔から細胞質へ放出します。このIP3Rは、さまざまな種の生物に普遍的に存在し、受精、発生、記憶や学習といった生物の生存に必須な現象において重要な役割を果たしている一方、細胞死を誘導することが定説として知られていました。
IP3Rは、分子量1.2MDa※2にも及ぶ巨大なタンパク質で、4量体※3を形成することでカルシウム放出チャネルとして機能します(図1)。IP3RにはIP3R1・IP3R2・IP3R3の3種のサブタイプが存在しますが、それらの細胞内局在や組織分布は必ずしも同じではなく、それぞれのサブタイプが生体内で特有の役割を果たしていると考えられています。中でも1型IP3受容体(IP3R1)は、脳で高い発現を示し、その機能を遺伝的に喪失したマウスが運動機能障害を引き起こすことが分かっています。しかし、IP3R1の脳における機能やその制御機構は十分には明らかとなっていません。また小胞体ストレスは、細胞内カルシウム恒常性を攪(かく)乱し、神経細胞死を誘導することが報告されていますが、IP3R1がそれらに関与するかどうかは明らかとなっていませんでした。研究チームは、このIP3R1に着目し、小胞体ストレスと細胞死との関係を解明することに取り組みました。
研究手法と成果
(1)小胞体ストレスによる神経細胞死へのIP3R1の関与
まず、研究チームは、小胞体ストレスが及ぼすIP3R1の機能への影響をカルシウムイメージング法※4で調べました。培養細胞(マウス神経芽細胞腫N1E-115細胞、ヒト子宮頸がん由来HeLa細胞)やマウス脳由来の神経細胞を、小胞体ストレスを誘導する薬剤(ツニカマイシン、タプシガルジン、ジチオスレイトール)で処理したところ、IP3R1のカルシウム放出活性が顕著に低下することが分かりました。
次に、IP3R1のカルシウム放出活性の低下が及ぼす細胞死への影響を調べるために、RNA干渉※5法によってIP3R1の発現を抑制した培養細胞(HeLa細胞)を用いて実験を行いました。驚くべきことに、長期間の小胞体ストレス条件下では、IP3R1の発現を抑制していない細胞に比べ、IP3R1の発現を抑制した細胞で細胞死(アポトーシス)が顕著に高まりました。この結果は、IP3Rが細胞死を誘導するという定説に反して、IP3R1が小胞体ストレスから細胞を守る働きをしていることを示唆しています。
この可能性を確かめるため、さらに、IP3R1の欠損したマウスを用いて実験を行いました。対照となる野生型マウスとIP3R1欠損マウスのそれぞれに、小胞体ストレスを誘導する薬剤(ツニカマイシン)を腹腔内投与したところ、IP3R1欠損マウスの小脳※6プルキンエ細胞※7において神経細胞死が誘導されました(図2)。
これらの結果から、小胞体ストレスが生じると、IP3R1の機能低下、つまり、カルシウム放出活性の低下が起こり、神経細胞死が誘導されることを示唆できました。
(2)小胞体ストレスによるIP3R1の機能低下の分子メカニズムの解明
IP3R1は、小胞体膜によって細胞質側の領域と小胞体内腔側 (L1、L2,、L3)の領域に分けられており(図3)、小胞体内腔側が小胞体ストレスを感知する領域として働くという仮説を立てました。特に、小胞体内腔側のL3領域は、L3VとL3C領域に分けることができ、非常に重要な特徴として、L3Cはチャネルポア(カルシウムが小胞体膜を通過する際の通り道)を形成します(図3)。さらに、このポア領域L3Cに隣接するL3Vは、IP3Rのサブタイプ間でアミノ酸配列が非常に低い相同性を示しており、サブタイプ特異的な機能に関与することが分かっています。通常、小胞体内腔は酸化状態を好むのに対し、小胞体ストレスでは、還元状態に傾くことが知られています。研究チームは、小胞体ストレス依存的にIP3R1への結合が変化するタンパク質が存在すると考え、IP3R1-L3Vの結合タンパク質の検索を行いました。その結果、酸化条件下で結合するタンパク質で、小胞体内腔に存在し、ストレスに応答して発現が誘導される分子シャペロン※8として知られる、GRP78を同定しました(図3)。
このGRP78と、IP3R1、IP3R2、IP3R3の3種のサブタイプの精製タンパク質を用いて結合を確認したところ、GRP78はIP3R1だけにL3Vを介して結合することを見いだしました。同様に、細胞内でもIP3R1とGRP78が結合すること、さらに小胞体ストレス下では、その結合が弱まることを明らかにしました。また、マウス脳内においてIP3R1とGRP78の局在が同じであることを確かめました(図4)。
次に、IP3R1のカルシウム放出機能に対するGRP78の役割を調べました。RNA干渉法によってGRP78の発現を抑制した培養細胞(HeLa細胞、N1E-115細胞)では、IP3R1のカルシウム放出活性が顕著に低下しました(図5)。一方、GRP78を過剰発現した神経細胞では、IP3R1のカルシウム放出活性が顕著に高まりました。この結果は、IP3R1がカルシウムチャネルとして機能するためには、GRP78が必要であることを示しています。
さらに、生化学的手法(ゲルろ過クロマトグラフィー※9、ショ糖遠心密度勾配遠心法※10)を用いて、GRP78がどのようにしてIP3R1を制御しているかを調べました。その結果、RNA干渉法によってGRP78の発現を抑制すると、IP3R1の4量体形成が阻害されることが分かりました。小胞体ストレス下においても同様に、IP3R1とGRP78の結合が低下し、IP3R1の4量体形成が阻害されることを見いだしました。これらの結果は、ストレスのない状態では、IP3R1がGRP78によって4量体形成の制御を受けていますが、小胞体ストレス下ではその相互作用が抑制され、カルシウムチャネルとして機能することができなくなることを示しています(図6)。また、この機能的な相互作用の低下は、ハンチントン病※11のモデルマウスでも観察され、IP3R1の機能低下と神経変性疾患の関与が示されました。
今後の期待
研究チームは、小胞体ストレスによるIP3R1の機能破壊が神経細胞死を誘導し、脳障害を引き起こすことを見いだしたことから、IP3R1がストレスから脳を守る働きをしていることを示しました。さらに、その分子メカニズムとして、通常状態ではGRP78というタンパク質によって4量体に組み立てられるIP3R1が、ストレス条件下では4量体を形成できなくなることを見いだしました。この発見は、脳を形成する神経細胞をストレスから守る機構が存在することを示した点で、大変大きな意義を持ちます。今後、学術的な貢献にとどまらず、ストレスに起因する神経疾患の発症機構の理解や治療法の確立を通して、医学・健康科学に大きく貢献することが期待されます。
発表者
理化学研究所
脳科学総合研究センター 疾患メカニズムコア 発生神経生物研究チーム
チームリーダー 御子柴 克彦(みこしば かつひこ)
Tel: 048-467-9745 / Fax: 048-467-9744
お問い合わせ先
脳科学研究推進部 企画課Tel: 048-467-9757 / Fax: 048-462-4914
(JSTの事業に関すること)
独立行政法人科学技術振興機構
イノベーション推進本部 研究プロジェクト推進部
小林 正(こばやし ただし)
Tel: 03-3512-3528 / Fax: 03-3222-2068
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.細胞死
細胞は、物理的なダメージや毒物などによる強い障害で死滅するだけでなく、増殖シグナルの欠如あるいはさまざまな刺激の結果、細胞の「自殺スイッチ」が入り細胞死を起こすことが知られている。このような細胞死は、プログラム細胞死またはアポトーシスと呼ばれる。 - 2.Da(ダルトン)
分子や原子の質量を表す単位。炭素の同位元素12C(炭素)原子の1個の質量を12Daとする。従って、1Da=1.661x10-27kg。一般には、1molあたりのタンパク質の相対質量である分子量の単位として便宜的に使用している。1MD=1×106Da - 3.4量体
分子やポリペプチド鎖などの単量体が4個重合、または会合したものをいう。IP3Rは4量体を形成することで、そのカルシウム放出機能を発揮することができる。 - 4.カルシウムイメージング法
カルシウムイオンと結合すると、蛍光を発したり、蛍光波長が変化するカルシウム蛍光指示薬を用いて、生体内のカルシウムイオン濃度の変化を測定する方法。 - 5.RNA干渉
2本鎖RNAが,相同な塩基配列をもつメッセンジャーRNAと特異的に対合し,切断することによって,遺伝子の発現を抑えてしまう現象。この現象を利用して人工的に2本鎖RNAを導入することで、人為的に任意の遺伝子の発現を抑制する手法がRNA干渉法。2006年、アンドリュー・ファイアーとクレイグ・メローはRNA干渉の発見の功績よりノーベル生理学・医学賞を受賞した。 - 6.小脳
脳幹に存在する運動中枢で、ほ乳類の脳では大脳に次いで大きな脳組織。末梢からの姿勢、筋肉活動、バランス、視覚情報などの入力を受け、大脳と連携しながら、スムーズな運動の制御、並びにその学習に中心的な役割を果たす。 - 7.プルキンエ細胞
小脳皮質の情報処理の中心的なニューロン。大型の特徴のある神経細胞で、非常に広範囲の神経情報を受け取り、統合することが知られている。小脳皮質のプルキンエ細胞層にだけ存在し、樹上突起を分子層に伸ばし、その軸索は白質を通って深部小脳核へ投射する。樹上突起では、入力神経線維である苔状線維と登上線維からの情報を受ける。 - 8.分子シャペロン
タンパク質は正常に働くためには正しく折り畳まれる必要がある。これを助ける一群のタンパク質が分子シャペロンと呼ばれ、熱ショックなどで誘導される。また分子シャペロンは、タンパク質の正確な折り畳みを助けるだけでなく、凝集や分解の防止や、タンパク質の活性制御にも関与している。 - 9.ゲルろ過クロマトグラフィー
カラムにつめた担体(さまざまな物質と接したときに化学的に反応せず、また試料と吸着しにくい球状の多孔性粒子)に試料を通して、分子を大きさの違いによって分離する手法。試料は担体の間をすり抜けて分子は溶出されるが、担体には小さな孔が開いており、小さい分子はその孔に入り込んで寄り道するため、大きい分子より遅く溶出される。 - 10.ショ糖遠心密度勾配遠心法
遠心チューブの底部から上部に向けて、次第に濃度が低下するようにショ糖溶液の密度勾配を作り、最上部に試料を重層して遠心することで試料中に含まれる物質を大きさや重さに応じて分離分画する方法。 - 11.ハンチントン病
主に中年以降に発症する常染色体優性遺伝の遺伝性慢性進行性の疾患。その臨床症状は主に不随意運動であり、知能障害、精神症状を伴う。主に、線条体の障害により不随意運動を生ずると考えられている。1983年に、ハンチントン病遺伝子が第4染色体短腕(4p16.3)に存在することが示され、1993年に、その原因遺伝子(ハンチンチン遺伝子)が同定された。
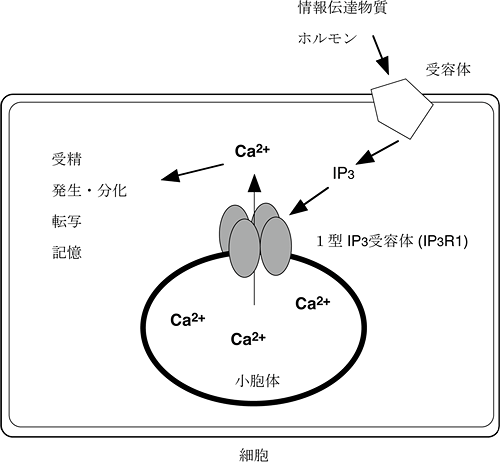
図1 IP3受容体の構造と機能
IP3受容体は、4量体を形成することでカルシウム放出チャネルとして機能する。細胞外刺激で産生されるIP3によって活性化され、カルシウム放出することで、さまざまな細胞応答に影響を及ぼす。
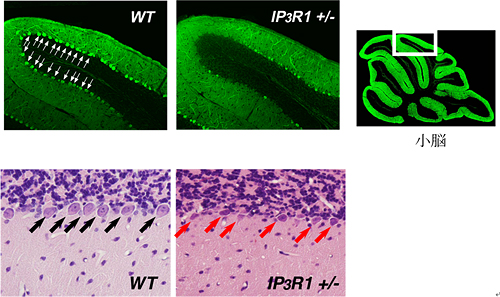
図2 IP3R1の欠損は、小胞体ストレスにおいて神経細胞死を誘導する
対照となるマウス(WT)とIP3R1の欠損したマウス(IP3R1 +/-)に小胞体ストレスを誘導する薬剤を投与したところ、IP3R1欠損マウスの小脳プルキンエ細胞において神経細胞死(脱落)が誘導された。写真は、カルビンディン染色(上段)、ヘマトキシリン・エオシン染色(下段)の結果。
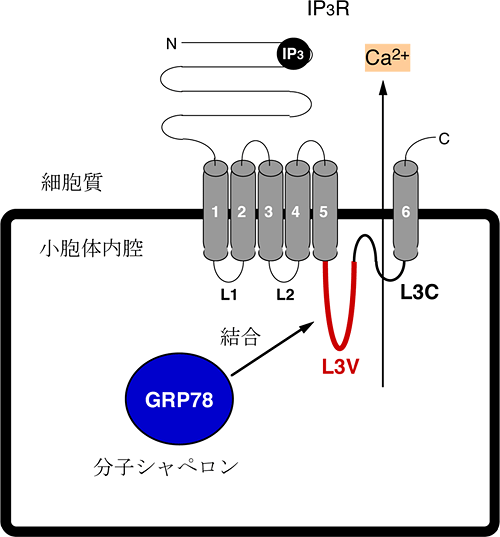
図3 IP3R1に小胞体内腔から結合するタンパク質GRP78の発見
IP3R1は、小胞体膜を6回貫通するタンパク質で、細胞質側と小胞体内腔側の領域に大別できる。細胞質側のIP3結合領域にIP3が結合するとポア領域L3Cを介してカルシウムが小胞体内腔から細胞質へ放出される。GRP78は、通常の小胞体内腔環境において、IP3R1のL3Cに隣接するL3V領域に結合するタンパク質として同定された。
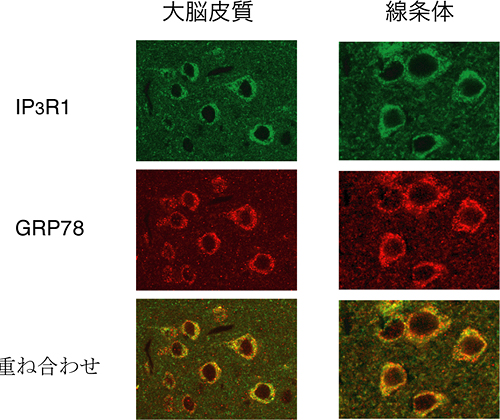
図4 マウス脳内でのIP3R1とGRP78の局在の様子
IP3R1とGRP78は、マウス脳内で同一の局在を示す。
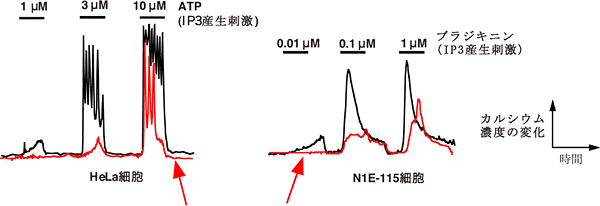
図5 GRP78の発現とIP3R1のカルシウム放出活性
RNA干渉法によるGRP78の発現抑制は、IP3R1のカルシウム放出活性を低下させる。
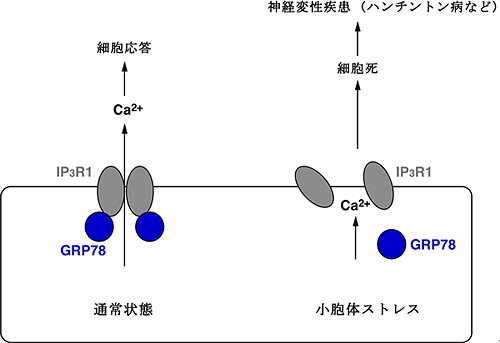
図6 小胞体ストレスによるIP3R1の機能低下の分子メカニズム
通常状態では、タンパク質GRP78によってIP3R1は4量体に組み立てられ、カルシウムチャネルとして機能することで、さまざまな細胞応答を制御することができる。一方、小胞体ストレス条件下では、GRP78がIP3R1に結合できないため、IP3R1は4量体を形成することができず、その結果、細胞死を誘導し、脳障害を引き起こすことになる。