2009年3月13日
理化学研究所
脂質メタボローム解析でスルホ脂質生合成の鍵となる新規遺伝子を発見
-スルホ脂質生合成を制御し、リン欠乏耐性植物や有用物質産生植物の生産へ新たな道-
ポイント
- 高性能質量分析計を駆使した植物脂質メタボロームの解析パイプラインを確立
- 新規遺伝子「UGP3」は、スルホ脂質生合成の初期段階で特異的に働く
- UGP3が関与するスルホ脂質生合成メカニズムは、イネやポプラなど植物全般に
要旨
独立行政法人理化学研究所(野依良治理事長)と国立大学法人東京工業大学(伊賀健一学長)は、植物に広く存在するスルホ脂質生合成に関与する新規遺伝子を発見し、その機能の同定に成功しました。これは理研植物科学研究センター(篠崎一雄センター長)メタボローム基盤研究グループの斉藤和季グループディレクター、岡咲洋三特別研究員らと東京工業大学の太田啓之教授、下嶋美恵特任助教らの研究グループによる共同研究の成果です。
肥料の三大要素に数えられるリンは、植物の生育に必須な元素の一つです。植物は、リンが欠乏すると、細胞膜を構成している主要脂質のリン脂質が減少し、それを補うように、スルホ脂質(硫酸化糖脂質)※1が増加して、膜機能を維持し生長を助けます。これまで、スルホ脂質の生合成過程の、最後の2段階に関与する2種類の遺伝子が特定されていましたが、この2つの遺伝子だけでは、スルホ脂質の生合成が十分に説明できないことから、さらに必要な因子が存在すると考えられていました。
研究グループは、まず高性能質量分析計※2を用いて、包括的な脂質分析(脂質メタボローム※3解析)のパイプラインを確立しました。次に、遺伝子共発現解析※4により候補遺伝子を絞り込み、シロイヌナズナでこの遺伝子の欠損変異体の脂質メタボローム解析を行って、スルホ脂質生合成の第一段階に関与し、律速酵素※5と考えられる酵素の遺伝子を同定しました。さらに、この遺伝子が、スルホ脂質の前駆体であるUDP-グルコース※6の葉緑体内での合成に関与することを明らかにしました。また、比較ゲノム解析によって、この遺伝子と類似性の高い遺伝子を、イネなどほかの植物からも発見しました。スルホ脂質は、多くの植物に普遍的に存在していることから、同様の生合成メカニズムが植物全般に存在することが示唆されました。
植物は、生育のためにリン脂質を含めて、さまざまなリン化合物を必要とします。しかし、肥料として使用されるリン鉱石は、近年枯渇が危惧されており、農業の将来にとって大きな問題となっています。発見した遺伝子を利用することで、スルホ脂質生合成の制御が可能になり、将来危惧されるリン欠乏に耐性な植物を作出できる可能性が出てきました。また、スルホ脂質は、抗がん作用、免疫抑制、抗エイズウィルス、抗炎症など、創薬につながる可能性のあるいくつかの有用な生理活性を持つことから、今回の成果により、植物を用いた有用物質の効率的な生産も期待されます。
本研究成果の一部は、JST戦略的創造研究推進事業(CREST)の研究課題「植物アミノ酸代謝のオミクス統合解析による解明」(研究代表者:平井優美 理研植物科学研究センター 代謝システム解析チーム チームリーダー)の一環として行ったもので、米国の科学雑誌『The Plant Cell』オンライン版(3月13日付け)に掲載されます。
背景
植物細胞の生体膜は、主にリン脂質や糖脂質から構成され(図1)、これらにはスルホ脂質(sulfolipid)と呼ばれる植物に特異的な脂質が含まれます(図2)。スルホ脂質の植物体中での含量は、生育環境によって変動し、特に植物の生育に必須な元素であるリンが少ない環境で育てると、リン脂質が減少し、それを補う形でスルホ脂質が顕著に増加します。また、スルホ脂質を作ることができない植物の変異体は、リンの含量が低い環境では生育が悪くなることも知られています。このため、スルホ脂質は、植物の劣悪な環境での生育に重要な役割を果たす脂質であると考えられています。これまで、スルホ脂質の生合成の最後の2つの段階に関与する遺伝子として、それぞれSQD1(UDP-スルホキノボース合成酵素遺伝子)とSQD2(スルホ脂質合成酵素遺伝子)が特定されています。しかし、この2つの遺伝子だけではスルホ脂質の生合成が十分に説明できないことから、さらに必要な因子が存在すると考えられていました。
研究手法と成果
- 1.スルホ脂質の合成に関与する新規遺伝子「UGP3」の同定
研究グループは、植物メタボロミクス研究の一環として、高性能質量分析計を用いた包括的な脂質分析(脂質メタボローム解析)のための解析パイプラインを構築しました。この解析パイプラインを用いて、スルホ脂質生合成に関与すると予想される遺伝子(候補遺伝子)の脂質メタボローム解析を行うことにしました。
候補遺伝子は、公共のDNAマイクロアレイ※7に基づく網羅的な遺伝子発現データを基に、シロイヌナズナの遺伝子発現パターンを解析し(遺伝子共発現解析)、2万種を超える遺伝子の中から3種類まで絞り込みました。
次に、候補遺伝子の欠損変異体をシロイヌナズナで作製し、解析パイプラインを用いた脂質メタボローム解析を行いました。その結果、スルホ脂質を特異的に蓄積しない変異体を見いだし、スルホ脂質の合成に関与する新たな遺伝子「UGP3」を同定しました(図3)。大腸菌でUGP3遺伝子を発現させ、このタンパク質の作用で生成する化合物を高性能質量分析計で解析した結果、UGP3遺伝子は、UDP-グルコース合成酵素遺伝子であることが分かりました(図4)。さらに、このUDP-グルコース合成酵素が細胞内のどこに輸送されるのかを追跡した結果、植物細胞内に存在する葉緑体に局在することが分かりました(図5)。 - 2.UGP3遺伝子の機能の解析
スルホ脂質には、硫酸化糖が構造の一部として結合しています。従って、UGP3遺伝子は、スルホ脂質の合成の第一段階として、硫酸化糖の前駆体であるUDP-グルコースを葉緑体中で合成する反応に関与することが推定されました。植物には、抗酸化物質であるフラボノイドのような物質が含まれており、これらの多くは配糖体※8として存在しています。UDP-グルコースは、これらの配糖体の主要な前駆体であるため、UGP3遺伝子を破壊した植物では、さまざまな配糖体の蓄積量にも変化が生じることが予想されました。しかし、UGP3変異体では、スルホ脂質以外の配糖体の蓄積量には大きな変動は認められませんでした。これらの結果から、この遺伝子はスルホ脂質の生合成に高度に特化した遺伝子であることが明らかとなりました(図5)。
UGP3遺伝子は、ほかの既知の遺伝子と非常に低い相同性しか示さなかったため、これまで知られていなかった新しい種類の遺伝子であるといえます。一方、比較ゲノム解析によって、イネ、ブドウ、ポプラ、コケ、シダなどの幅広い植物で進化的に保存されている遺伝子であることも明らかになりました。スルホ脂質は、多くの植物に普遍的に存在していることから、植物全般に同様の生合成メカニズムが存在することが示唆されました。
今後の期待
スルホ脂質の生合成の最後の2段階に関与する遺伝子は既に特定されていましたが、これらの遺伝子の発現レベルを改変することで、スルホ脂質の蓄積レベルが向上した例はありませんでした。今回発見した遺伝子は、代謝系路において上流の反応に関与しているため、スルホ脂質生合成に必要な前駆体のレベルをより直接的に制御できることが期待されます。また、この遺伝子は、生合成の律速段階に関与している可能性があります。従って、今回発見した遺伝子を使うことで、植物体中でのスルホ脂質生合成経路の制御がより容易となると期待されます。
スルホ脂質の植物体中での機能に関しては、まだ不明な点もあります。葉緑体に局在するいくつかのタンパク質には、一定の割合でスルホ脂質が結合していることが知られていますが、それらの詳細な役割についてはよく分かっていません。今回の発見は、植物中でのスルホ脂質が果たす役割を解明する上で、新たな切り口を与えてくれることも期待されます。
植物は、リン脂質を含めてさまざまなリン化合物を必要としており、リンは植物の生育にとって必須の元素です。しかし、肥料として使用されるリン鉱石は近年枯渇が危惧されており、農業の将来にとって大きな問題となっています。今回発見した遺伝子を利用することでスルホ脂質生合成の制御が可能になり、将来危惧されるリン欠乏に耐性な植物を作出できる可能性があります。さらに、スルホ脂質は、抗がん作用、免疫抑制、抗エイズウィルス、抗炎症など、創薬につながる可能性のあるいくつかの有用な生理活性も持っています。従って、今回の成果は、植物を用いた有用物質の効率的な生産を可能にすることも期待されます。
発表者
理化学研究所
植物科学研究センター メタボローム基盤研究グループ
グループディレクター 斉藤和季(さいとう かずき)
特別研究員 岡咲洋三(おかざき ようぞう)
Tel : 045-503-9488 / Fax : 048-467-9489
お問い合わせ先
横浜研究推進部 企画課Tel : 045-503-9117 / Fax : 045-503-9113
国立大学法人東京工業大学 バイオ研究基盤支援総合センター
教授 太田啓之(おおた ひろゆき)
特任助教 下嶋美恵(しもじま みえ)
Tel : 045-924-5736 / Fax : 045-924-5823
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.スルホ脂質(硫酸化糖脂質)
スルホキノボースという硫黄を含むグルコース誘導体を部分構造とする膜脂質。通常、細胞膜などにはほとんど含まれておらず、主に葉緑体の膜に存在する。 - 2.高性能質量分析計
物質の分子量を高い精度で測定する機器。試料をイオン化し、化合物の質量電荷比(質量を電荷数で割った値)から物質を同定・定量する。高感度で物質を検出できるため、植物ホルモンのような微量物質の分析に有用。 - 3.メタボローム
細胞内に含まれる低分子代謝産物の総体を指す用語。代謝産物を包括的かつ網羅的に解析することにより、生体内での化学反応を全体的に把握可能にする技術を含め、ゲノム情報から転写産物mRNA情報(トランスクリプトーム)、タンパク質情報(プロテオーム)に続く代謝産物総体(メタボローム)についてのポストゲノム科学分野が、メタボロミクスと呼ばれる。 - 4.遺伝子共発現解析
複数の遺伝子の発現パターンの相関の程度(相関係数:+1から-1までの数値で表される)を基準として、機能が未同定の遺伝子群から目的の遺伝子を見つけ出す解析手法。多くの場合、同一の生合成経路に属する遺伝子群は協調的に発現することから、相関係数が高い遺伝子群は、同一もしくは近接する経路、あるいは同時期に起こっている反応に関連する可能性が高いと考えられる。 - 5.律速酵素
ある化合物の合成に関わる反応プロセスの中で、特別に遅い反応段階があると、ほかの反応がいくら速くても、その遅い段階の反応速度が全体としての速度を決定してしまう。これを律速段階といい、これを触媒する酵素を律速酵素という。 - 6.UDP-グルコース
グルコースの活性化した形で、代謝過程でグルコース転移酵素の基質となる。また、UDP-ガラクトースやUDP-グルクロン酸に変換されて、ガラクトースやグルクロン酸を含む多糖の原料になる。 - 7.DNAマイクロアレイ
細胞内の遺伝子発現量を測定するために、多数のDNA断片をプラスチックやガラスなどの基板上に高密度に配置した分析器具のこと。 - 8.配糖体
天然の化合物(フラボノイド、テルペノイド、植物ホルモンなど)は糖(グルコース、ラムノース、キシロースなど)が結合した状態で存在することが少なくない。天然の化合物は配糖化により、親水性が増したり、安定化したり、不活性化することから、化合物の蓄積やその機能の制御に関与していると考えられている。
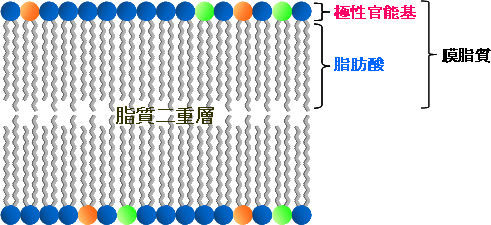
図1 生体膜の模式図
植物の細胞にはリン脂質や糖脂質のようなさまざまな膜脂質が存在し、それぞれの膜脂質には異なる極性官能基を結合している。これらの膜脂質は、生体中では脂肪酸同士が向かい合う形で二重層を構成し、その結果さまざまな生体膜が形成される。
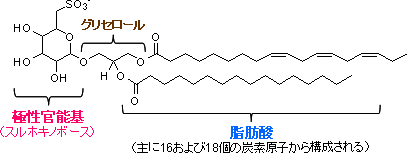
図2 スルホ脂質の構造
スルホ脂質はスルホキノボースとグリセロール、そして2種類の脂肪酸から構成される。生体中には異なる脂肪酸が結合したスルホ脂質が複数存在する。
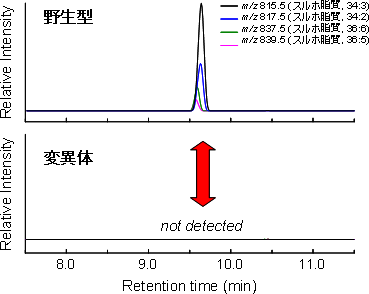
図3 野生型と変異体におけるスルホ脂質の蓄積パターンの比較
野生型からはスルホ脂質は検出されたが、UGP3遺伝子を欠損する変異体からはまったく検出されない。
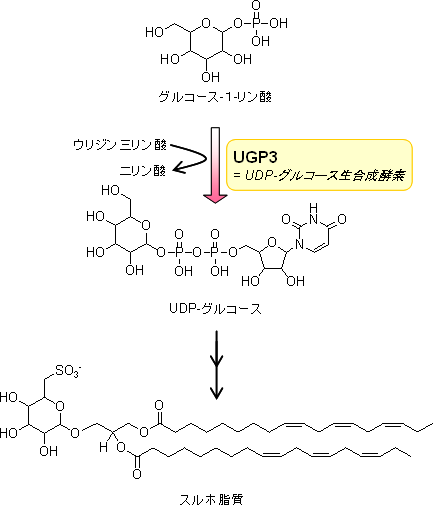
図4 発見した新たなスルホ脂質生合成機構
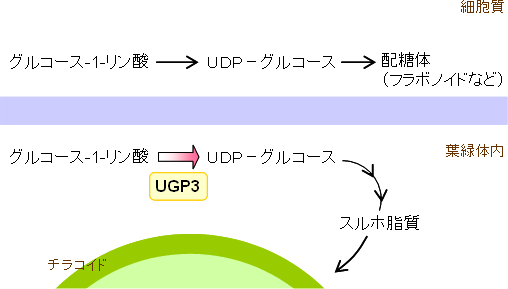
図5 UGP3の細胞内局在性とその機能的特異性
植物はスルホ脂質以外にもさまざまな配糖体(フラボノイド配糖体など)を合成するが、UGP3は葉緑体中に局在してスルホ脂質生合成に特異的に関与する。