2009年6月12日
独立行政法人 理化学研究所
独立行政法人 科学技術振興機構
抑制性神経伝達を制御する新たな分子機構を、量子ドットを活用し発見
-シナプスにおける受容体の側方拡散が、GABA作動性シナプス伝達効率を決める-
ポイント
- 興奮性神経活動に依存して、シナプス内のGABAA受容体の側方拡散が増加
- 細胞内カルシウム濃度が、GABAA受容体の側方拡散を制御
- てんかんや統合失調症などの治療法確立に貢献する、新たなターゲットを提示
要旨
独立行政法人理化学研究所(野依良治理事長)と独立行政法人科学技術振興機構(以下JST、北澤宏一理事長)は、抑制性神経伝達効率の制御に神経細胞膜上のGABAA受容体※1の側方拡散※2がかかわっていることを世界で初めて明らかにしました。これは、理研脳科学総合研究センター(利根川進センター長)発生神経生物研究チームの御子柴克彦チームリーダー(JST戦略的創造研究推進事業 発展研究:SORSTの「カルシウム振動プロジェクト」研究総括)、坂内博子基礎科学特別研究員、ローニー研究ユニットのトーマス・ローニーユニットリーダー(元・記憶学習機構研究チーム研究員)やフランス・パリ高等師範学校とINSERM(国立衛生医学研究所)U789のアントワーヌ・トリラーグループディレクターらによる共同研究の成果です。
神経細胞は、シナプス※3という特殊な構造を介して情報の伝達を行っています。このシナプス伝達効率の柔軟な変化「シナプス可塑性」は、記憶・学習の基礎となり、シナプス伝達の異常は脳神経疾患の原因となります。シナプスでは、前シナプスから放出される神経伝達物質がシナプス後膜※3に局在する神経伝達物質受容体に結合して信号が伝わります。このシナプスにおけるシナプス伝達効率を決めるファクターの1つは、シナプス後膜に局在する神経伝達物質受容体の数です。
研究グループは、抑制性神経伝達をつかさどるGABAA受容体に注目して、シナプス内の受容体数を制御する分子機構の解明に取り組みました。その結果、神経興奮が過剰になるとシナプス内のGABAA受容体数は減少するのに対し、細胞膜上のGABAA受容体の総数は変化しないことを見いだしました。また、量子ドット※4を用いて細胞膜上のGABAA受容体の動きを1分子レベルで追跡したところ、興奮性神経活動の増加に伴って受容体の側方拡散が増加し、シナプス後膜における受容体の安定性が著しく減少することが明らかとなりました。すなわち、神経の興奮が高まるとGABAA受容体が細胞膜上を動きやすくなり、シナプス内に留まることができなくなった結果、シナプス後膜の受容体数が減少していました。さらに、細胞内カルシウム濃度の上昇によるカルシニュリン※5の活性化が、受容体の側方拡散を増加させることが分かりました。これまで、シナプス伝達効率を決める鍵となっているシナプス内の受容体数は、細胞膜上の受容体総数に比例して増減すると考えられてきました。今回の発見は、シナプス伝達効率決定機構に対し、「側方拡散制御による受容体数の増減」という新しい分子機構を提唱するものです。本研究成果は、米国の科学雑誌『Neuron(ニューロン)』(6月11日号)に掲載され、表紙を飾ります。
背景
私たちの神経ネットワークにおける情報伝達は、神経細胞(ニューロン)を興奮させる興奮性シナプスと、興奮を抑える抑制性シナプスの絶妙なバランスの上に成り立っています。興奮性シナプスは、神経伝達のシグナルである活動電位を発生させるために必要不可欠ですが、抑制性シナプスも脳の正常な機能のために極めて重要な役割を担っています。例えば、抑制性シナプスの1つであるGABA作動性シナプスによる適所・適量な抑制性神経入力は、ほ乳類の新生児における視覚発達の基礎過程である臨界期の開始に必要とされています。また、GABAA受容体の異常は、てんかん、不安障害、ハンチントン病、アンジェルマン症候群、fragile X syndrome、統合失調病、薬物依存症など、さまざまな脳神経疾患を引き起こします。従って、これらの脳疾患治療のためにも、GABA作動性シナプスの制御機構を知ることは重要とされていますが、これまでGABA作動性シナプス制御にかかわる分子機構の実体は、ほとんど明らかになっていません。
GABA作動性シナプス伝達効率は、シナプス後膜に局在するGABAA受容体の数に依存しています。従来、細胞膜上に発現する受容体の総数がシナプス後膜の受容体数の決定因子であると考えられてきました。ところが近年になって、細胞膜上の神経伝達物質受容体が側方拡散によりシナプス内外をダイナミックに出入りしていることが明らかになってきました。この発見に伴い、シナプス後膜に局在する受容体の数を決定する新たな要因として「受容体の側方拡散」が考えられるようになりました。これまでに、脳の海馬※6という部位で神経細胞が過剰に興奮すると、GABA作動性シナプス伝達効率が弱くなる現象が報告されており、「抑制性シナプス可塑性」として知られています。この現象は記憶学習やてんかんの病態発現にかかわると考えられていますが、詳細な分子機構は解明されていませんでした。研究グループは、GABA作動性シナプス伝達効率がGABAA受容体の側方拡散制御を介して決定されるかどうかを、ラット海馬の培養神経細胞を材料として解明することに取り組みました。
研究手法と研究成果
(1)シナプスおよび細胞膜上のGABAA受容体数の変化
ラット海馬の培養神経細胞の興奮性を人為的に高めると、GABA作動性シナプス1個あたりの電気的応答が小さくなることが確認できました(図1)。免疫染色法を使って、シナプス後膜に局在するGABAA受容体数の変化を調べたところ、神経興奮の増加から5分以内に、GABAA受容体の数が50%程度に減少することが分かりました(図2A)。ウエスタンブロット法を用いて、細胞膜上に発現するGABAA受容体の総数を解析したところ、神経興奮が高まっても細胞表面のGABAA受容体数は、まったく変化しませんでした(図2B)。この結果は、シナプス後膜に局在するGABAA受容体数が減ったのは、細胞膜上に発現している受容体が減少したからではなく、シナプス後膜に密集していたGABAA受容体がシナプス外に移動したためであることを示しています。
(2)シナプスにおけるGABAA受容体の神経活動依存的な側方拡散の計測
GABAA受容体は、細胞膜上を2次元ブラウン運動により側方拡散しています。側方拡散は、量子ドットを用いた1分子イメージング法を用いると、数十nm(1nmは10-9m)という空間解像度で解析することができます。神経細胞膜上のGABAA受容体1分子を量子ドット1個で標識し(図3A)、蛍光顕微鏡下でその側方拡散が神経活動によって変化するか否かを検討しました(図3B)。その結果、過剰な興奮性神経活動により、GABAA受容体がシナプス内に滞在する時間が減少し、シナプスへの出入りの回数が増加することを見いだしました(図4)。また、シナプスにおけるGABAA受容体の拡散係数※7と、受容体が拡散できる領域の大きさも、神経活動依存的に増加していることが分かりました(図5)。これらの結果は、興奮性神経活動の増加がGABAA受容体の側方拡散が増加し、GABAA受容体をシナプス後膜から流出しやすくする現象を引き起こすことを意味しています。
(3)GABAA受容体の側方拡散を増加させる細胞内シグナルの探索
研究グループは、GABAA受容体の側方拡散が、細胞内カルシウム濃度の上昇に伴って増加していることを見いだしました(図6)。また、細胞内カルシウム濃度の上昇は細胞外からのカルシウム流入で生じ、それによって活性化したカルシニュリンという酵素の働きによってGABAA受容体の側方拡散が増加することを突き止めました。細胞外からのカルシウム流入とカルシニュリンの働きは、海馬の抑制性シナプス可塑性にも必要とされています。従って、カルシウム流入とカルシニュリンの活性化により、シナプス内でGABAA受容体の側方拡散が増加することが、抑制性シナプス可塑性を引き起こす分子機構であると考えられます。
今後の期待
これまでの研究では、シナプス伝達効率を決めるのは細胞膜上に発現する受容体の総数であると考えられてきました。それに対し本研究は、シナプス伝達効率の制御に「側方拡散制御を介したシナプス後膜に局在する受容体数の増減」という新たな分子機構が働いていることを提唱するものです(図7)。GABA作動性シナプスの異常は、てんかん、統合失調症をはじめとする多くの脳神経疾患の原因であり、GABA作動性シナプス伝達効率の出産や加齢による変化が、産後のうつ病やアルツハイマー病の記憶障害に関与することが知られています。今回の成果は、神経興奮が過剰になっているてんかん患者の脳でGABAA受容体の側方拡散が増加している可能性を世界で初めて示した点で、大きな意義があります。今後、GABAA受容体の側方拡散制御の分子機構をさらに詳しく解明することで、てんかんをはじめとするさまざまな脳神経疾患に対する治療法の確立に貢献することが期待できます。
発表者
理化学研究所
脳科学総合研究センター 発生神経生物研究チーム
チームリーダー 御子柴 克彦(みこしば かつひこ)
基礎科学特別研究員 坂内 博子(ばんない ひろこ)
Tel: 048-467-9745 / Fax: 048-467-4744
お問い合わせ先
脳科学研究推進部 納富 さより(のうどみ さより)
Tel: 048-467-9654 / Fax: 048-462-4914
(JSTの事業に関すること)
独立行政法人科学技術振興機構
イノベーション推進本部研究プロジェクト推進部
小林 正(こばやし ただし)
Tel: 03-3512-3528 / Fax: 03-3222-2068
報道担当
独立行政法人理化学研究所 広報室 報道担当Tel:048-467-9272 / Fax:048-462-4715
独立行政法人科学技術振興機構 広報ポータル部
Tel: 03-5214-8404 / Fax: 03-5214-8432
補足説明
- 1.GABAA受容体
中枢神経系において速い抑制性神経伝達を担う、5つのサブユニットから構成されるイオンチャネル。神経伝達物質の1つのγアミノ酪酸(GABA)が結合することで開口し、塩化物イオン(Cl-)を透過させる。 - 2.側方拡散
1972年にSingerとNicolsonにより提唱された生体膜モデルによると、細胞膜は脂質二重層とモザイク状に入り混じったタンパク質により構成されている。これらの細胞膜構成要素は流体としての性質を持ち、細胞膜の中をブラウン運動している。この2次元的なブラウン運動を、側方拡散と呼ぶ。 - 3.シナプス、シナプス後膜
神経細胞同士の情報伝達にかかわる構造。情報を伝える細胞と伝えられる細胞の間には約20nmのすき間がある。情報を伝える細胞はこのすき間に神経伝達物質を放出し、伝えられる細胞側の神経伝達物質受容体がそれを受けとることにより神経情報が伝わる。情報を受け取る細胞のシナプスの細胞膜には神経伝達物質が密集しており、この構造を特に「シナプス後膜」と呼ぶ。 - 4.量子ドット
直径約15~25nmの半導体素材からなるナノ結晶。生命科学の分野では蛍光プローブとして用いる。従来の蛍光色素と比べてシグナルが強く、退色しにくいという利点があり、1分子イメージングに適している。また粒子の大きさによって様々な波長の蛍光を発することから,複数の分子を同時にイメージングする際の強力なツールとなっている。 - 5.カルシニュリン
カルシウムによって活性化されるタンパク質脱リン酸化酵素。 - 6.海馬
記憶の形成に不可欠な脳の部位。 - 7.拡散係数
側方拡散の速さを表す物理量。この値が大きいほど、分子の拡散は速くなる。
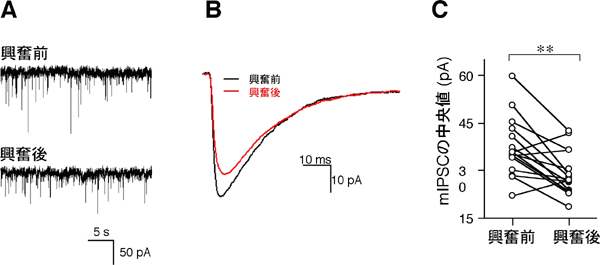
図1 過剰に神経が興奮すると、GABA作動性シナプスの電気的応答は小さくなる
A:海馬培養神経細胞におけるGABA性微小抑制性後シナプス電流(mIPSC)記録の一例。下向きのスパイクが、シナプスの電気的応答を反映している。
B:Aで記録されたmIPSC175個を平均化したもの。興奮後(赤)は、興奮前(黒)に比べてGABA作動性シナプス応答が小さくなった。
C:16個の神経細胞で、興奮を与える前後のmIPSCの中央値を比較した。過剰な神経興奮により、mIPSCは有意に低下する。16個中11個の細胞で、興奮後のmIPSCは興奮前に比べて20%以上減少していた。**は統計的に差が有意であることを示している。
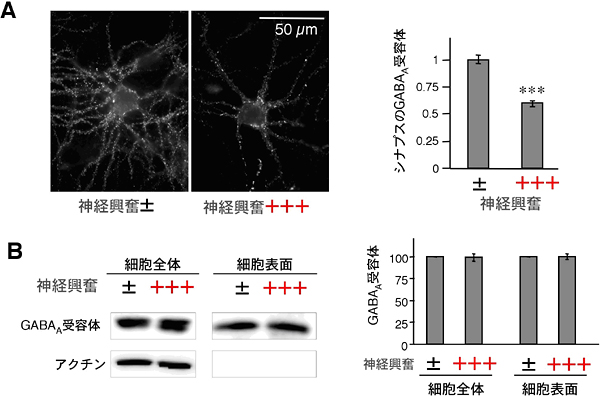
図2 神経が興奮するとシナプスのGABAA受容体数が減るが、細胞表面の受容体数は変わらない
A:海馬培養神経細胞のGABAA受容体を免疫染色した像(左)。5分間過剰な神経興奮が続いた細胞(神経興奮+++)では、免疫染色のシグナルが低下している。シナプスのGABAA受容体シグナルを定量化したところ(右のグラフ)、過剰に神経興奮した細胞(+++)のシグナル強度は興奮していない細胞(±)の約50%まで減少していた。***は統計的に差が有意であることを示している。
B:細胞全体と細胞表面のGABAA受容体のタンパク質量をあらわすウエスタンブロットの結果(左)。アクチンは細胞内にしか存在しないタンパク質なので、細胞表面にはシグナルがない。GABAA受容体のシグナルを定量したところ(右)、細胞全体,細胞表面ともに、過剰に神経興奮してもGABAA受容体のタンパク質量は減らないことが明らかとなった。
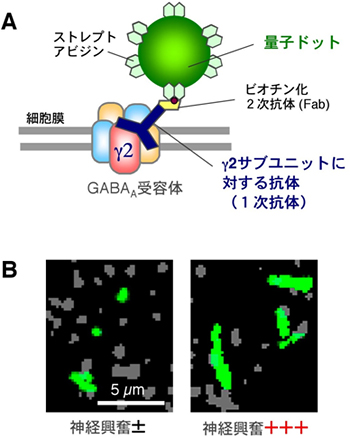
図3 量子ドット1分子イメージングによるGABAA受容体側方拡散の解析
A:量子ドットで細胞表面のGABAA受容体を1分子標識する方法。1次抗体、ビオチン化2次抗体を介して、ストレプトアビジン融合量子ドットで標識する。GABAA受容体は5つのサブユニットから構成されているが、その中で1つしか存在しないγ2サブユニットに対する抗体を1次抗体として用いることで、受容体と量子ドットを1:1で標識できる。量子ドットの動きは蛍光顕微鏡で追跡する。量子ドット1分子イメージング法では内在性分子の側方拡散を観察できるという点で、外来性蛍光タンパク質を用いた従来の生細胞イメージング法に比べて優れている。
B:量子ドットで標識されたGABAA受容体が38.4秒間に動いた範囲(緑)。グレーはFM4-64という蛍光色素で標識したシナプス。神経興奮が増加すると(+++)、同じ時間内に動く範囲が広くなる。すなわち受容体の側方拡散が増加している。
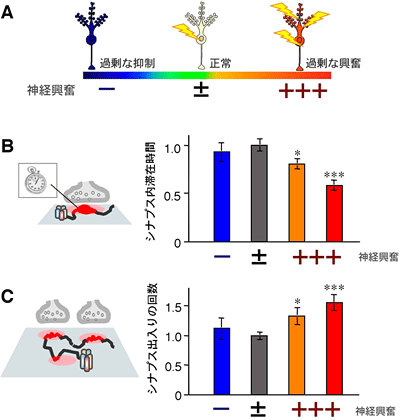
図4 神経興奮が過剰になると、GABAA受容体のシナプス内滞在時間が減少し、シナプスへの出入りの回数が増える
A:薬剤を用いて海馬培養神経細胞の神経活動を人工的に変化させ、抑制過剰状態(-)、興奮過剰状態(+++)を作り出した。正常(±)は薬剤未処理の状態。
B:さまざまな興奮状態の神経細胞において、量子ドットで標識されたGABAA受容体1個のシナプス内滞在時間を測定したところ、神経興奮が過剰になると(+++)シナプス内滞在時間が減少した。
C:単位時間あたりのシナプスへのGABAA受容体の出入りの回数。興奮過剰になると(+++)シナプスへの出入りの回数が増加する。
B、Cのグラフの値は、正常状態(±)の平均値を1.0として正規化した値を示す。
*、***は統計的に差が有意であることを示し、*の数が多いほど有意差がある。
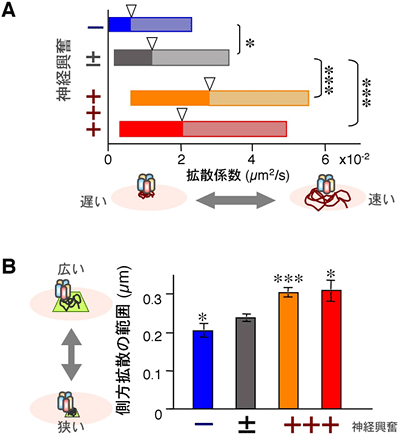
図5 GABAA受容体の側方拡散は、神経興奮依存的に増加する
A:側方拡散の速さの指標である拡散係数の解析。抑制過剰な状態(-)では、GABAA受容体の側方拡散は遅くなり、興奮過剰な状態(+++)では速くなることを示している。
B:シナプス内でGABAA受容体が側方拡散できる範囲を測定したところ、神経興奮が増加するにつれ、側方拡散できる範囲が拡大されることが分かった。
*、***は統計的に差が有意であることを示し、*の数が多いほど有意差がある。
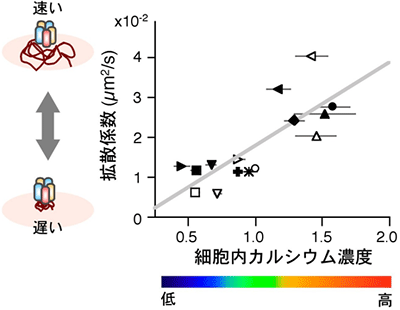
図6 GABAA受容体の側方拡散は細胞内カルシウム濃度に依存する
さまざまな薬剤処理条件下で測定された細胞内カルシウム濃度に対し、同条件におけるGABAA受容体の拡散係数の中央値をプロットしたグラフ。細胞内カルシウム濃度は、カルシウム指示薬fluo-4の蛍光強度を、薬剤刺激前の値を1.0として正規化した値の平均値として示している(エラーバーは標準誤差)。細胞内カルシウム濃度が高くなると、側方拡散が速くなっていることが分かる。
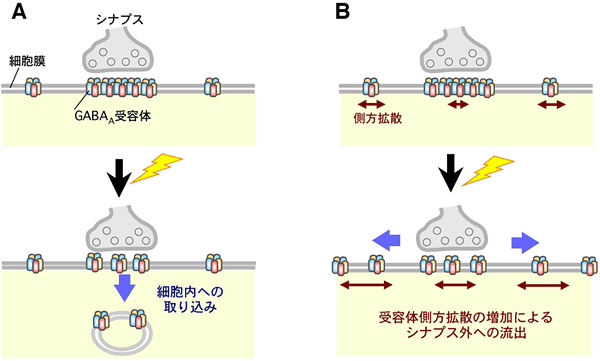
図7 本研究の結果から推測される、抑制性シナプス可塑性の新しい分子機構
A:これまで考えられてきた抑制性シナプスの分子機構は、GABAA受容体が細胞膜表面から細胞内に取り込まれるため、シナプスに局在できる受容体数が少なくなるというものであった。実際、興奮性シナプスにおいては、グルタミン酸受容体の一種であるAMPA受容体の細胞内への取り込みが神経伝達効率の減少を引き起こす原因となっている。
B:今回提唱する抑制性シナプスの分子機構は、側方拡散が増加することによりシナプス内に密集していたGABAA受容体がシナプス外に流出するため、抑制性シナプス伝達効率が低下するというものである。この新しい分子機構は、抑制性シナプス可塑性誘導に必要不可欠な、最初のステップであると考えられる。また、この現象は、神経興奮が過剰になっているてんかん患者の脳で実際に起こっていると予想される。