2012年10月19日
独立行政法人理化学研究所
単純な生化学反応から自律振動子を作る仕組みを解明
-タンパク質で構成された、細胞内で働く「人工時計」の設計指針に-
ポイント
- 2つの酵素と1つの基質による周期的な修飾状態の変動で自律振動子が生成
- 多数の基質1分子振動子が同期し巨視的な振動が生成する設計原理を提案
- 未知の細胞内自律振動子を見つける手がかりになると期待
要旨
理化学研究所(野依良治理事長)は、リン酸化/脱リン酸化といった可逆的なタンパク質の修飾から自律振動子※1を作る仕組みの一端を解明しました。これは、理研生命システム研究センター(柳田敏雄センター長)合成生物学研究グループの上田泰己グループディレクター、大出晃士特別研究員、理研発生・再生科学研究センター(竹市雅俊センター長)システムバイオロジー研究プロジェクト(上田泰己プロジェクトリーダー)のジョリー クレィグ チャールズ(Craig C. Jolley)国際特別研究員らの研究グループによる成果です。
朝に目覚めて夜眠くなるように、私たち生物の多くの機能は周期的に働いています。このような周期的な振る舞い(振動)を作る元になるシステムを「自律振動子」と呼んでいます。自律振動子を作って「人工時計」として働かせることができれば、細胞の周期的な振る舞いを自由に操れる可能性があります。しかし、自律振動子の具体的な仕組みや作成方法はまだ、よく分っていませんでした。
研究グループは、一般的なタンパク質の可逆的修飾機構※2だけで自律振動子が生成できるのではないかと考えました。そこで、最も良く知られたタンパク質による化学修飾の酵素反応の速度を表す反応式「ミカエリス・メンテン式」※3を用いて、タンパク質の修飾状態が時間とともに振動する条件をコンピューターシミュレーションによって探索しました。その結果、周期的な修飾状態の振動を作るために必要な最少セットとして、「複数箇所に修飾を受ける基質タンパク質」、「修飾を触媒する酵素」、「脱修飾を触媒する酵素」の3種類のタンパク質があれば振動子を形成できることを発見しました。さらに、自律振動子を形成するためには酵素と基質の結合の強さ(親和性)と、修飾/脱修飾の反応速度に特定の組み合わせパターンが必要であることが分かりました。
今回、細胞内で自律振動子として働く酵素や基質が満たすべき条件を明らかにしました。この成果は、未知の細胞内自律振動子を見つける手がかりになるだけでなく、人工的にタンパク質の自律振動子を作り出し、細胞に新たな機能を持たせる際の設計指針になります。細胞内で「人工時計」として働かせて、細胞の周期的な振る舞いを自在に操ることができると、体内時計の乱れが原因となる不眠症などのリズム疾患の治療などに役立つと期待できます。
本研究成果は、米国の科学雑誌『Cell Reports』オンライン版(10月18日付け:日本時間10月19日)に掲載されます。
背景
生物が示す多くの現象は周期性を持ちます。例えば、私たちが目を覚ましたり眠りに落ちたりするサイクルはおおよそ24時間周期で生じます。あるいは、カエルの受精卵はおおよそ30分ごとに細胞分裂を繰り返します。こうした周期的な振る舞い(振動)を引き起こす元となるシステムを「自律振動子」と呼びます。これまで細胞内でどのような因子がどのような仕組みで働き、自律振動子を形成しているのかについて、多くの研究が行われてきました。その中でも、私たちの1日周期で生じる生理現象に深く関わる概日時計は、最も理解が進んでおり、概日時計を駆動する遺伝子・タンパク質がさまざまな生物種で同定されています。それらがどのようにして自律振動子を形成するかについては、大きく分けて2つの仕組みが考えられています。
1つは、遺伝子の転写ネットワーク※4に基づくもので、転写因子の負のフィードバック制御※5によるものです。これについては、数理モデルによる定量的な理解が進んでおり、2011年に研究グループは、この仕組みが哺乳類の概日時計に重要な役割を果たすことを実際に細胞を用いた実験で示しました(2011年1月14日プレスリリース)。
2つ目は、タンパク質の可逆的な翻訳後修飾によるものです。これは、化学修飾を触媒する酵素と修飾を受ける基質の組み合わせにより、基質の修飾状態(基質が修飾されたり、脱修飾されたり)が時間とともに変動することで機能します。シアノバクテリアの概日時計※6は、この仕組みに基づいています。その機能を担うタンパク質は、多くの基質や酵素にはないいくつか特徴的な性質があります。そのため、一般的な酵素や基質を用いたタンパク質修飾に基づく自律振動子が作れるのか、また、そのために必要な最低条件が何かについては十分に理解されていませんでした。
研究グループは、一般的な酵素と基質の反応機構を用いた数理モデルを解析して、タンパク質修飾に基づく振動子を作るために必要な条件を検討しました。
研究手法と成果
研究グループはまず、自律振動子を形成するために最低限必要なタンパク質として、修飾酵素、脱修飾酵素、基質をそれぞれ1種類、計3種類のタンパク質だけを対象にしました。次に、酵素反応の速度を記述するとき、最も一般的に用いられている「ミカエリス・メンテン式」に従って修飾/脱修飾反応が起こると想定しました(図1)。この反応メカニズムでは、酵素と基質が結合する強さ(親和性)と基質が酵素によって修飾・脱修飾を受ける速さ(反応速度)の2つの要素で酵素反応の進行を表すことができます。そして、コンピューターを用いてこの2つの要素をさまざまに変更したときの基質修飾状態の時間変化をシミュレートしました。その結果、基質タンパク質の2カ所以上が修飾を受けると、修飾状態が振動する場合があることを発見しました(図2)。このとき、振動する場合と振動しない場合の違いを調べるため、12億通りの親和性と反応速度の組み合わせをコンピューターでシミュレーションし、どの組み合わせで振動が生まれるのかを調べました。その結果、振動を作る組み合わせには、2つの特徴があることが分かりました。1つは、対となる速度の速い修飾反応・脱修飾反応の方向の向きがそろっていること、2つ目は特定の修飾状態の基質が酵素と強く結合することです(図3)。この際、基質1つ1つの修飾状態に時間的な差が生じることあります。“遅れた修飾状態”の基質と酵素との親和性が高い場合には、“進んだ修飾状態”の基質が次の修飾状態になるために必要な酵素まで奪い取ることになります(図4)。そして、全ての基質が同じ修飾状態になってはじめて、全ての基質はいっせいに次の修飾状態に進むことができます。これにより、時間とともに修飾状態がいっせいに変動する自律振動子が形成されると考えられます。
実際に、遅れた基質が次の修飾状態に進む速度を遅くすると、基質集団の足並みがそろうまでに時間がかかり振動の周期が長くなることが分かりました(図5)。また、2つ目の特徴である酵素と基質の親和性を低くすると、基質集団の足並みが乱れ、基質集団の同調した振動は見られなくなり、自律振動子は形成されませんでした(図6)。これらから、特定の親和性の高さや反応速度を調整することによって、振動の振幅や周期長を制御できることが明らかとなりました。
すなわち可逆的なタンパク質の修飾から振動が生み出される原理は、基質1分子ごとが決まった順序で修飾を受けることで1分子振動子として働くこと、そして、それらの状態が基質間で揃うことで、同期した1分子振動子が基質集団としての修飾状態の自律振動を生み出すこと、ということが分かりました。
今後の期待
今回の成果で、一般的な修飾酵素、脱修飾酵素と複数の修飾箇所を持つ基質があると、振動子が形成可能なことが分かりました。興味深いことに、今回明らかになった振動の仕組みは、シアノバクテリア概日時計タンパク質で提唱されている仕組みとよく似ていました。基質の複数の場所が特定の酵素によって修飾を受ける例は概日時計に関わるタンパク質をはじめ、他の周期的な現象に関わるタンパク質においても数多く知られています。従って、タンパク質修飾に基づく振動子は、シアノバクテリア概日時計にみられるような特殊な例ではなく、他の生物種の多様な周期現象でも機能している可能性があります。
さらに、振動子形成に必要な結合の強さ、反応速度のパターンが明らかになったことで、振動子形成に適した酵素や基質の組み合わせを検討することができるようになりました。今後、酵素と基質の結合や、反応速度を予測し改変できるようになると、細胞内で人工的に設計したタンパク質振動子を「人工時計」として機能させ、細胞の周期的振る舞いを自在に操り、体内時計の乱れが原因となる不眠症などのリズム疾患の治療などに役立てることができると期待できます。
原論文情報
- Craig C. Jolley, Koji L. Ode, Hiroki R. Ueda.“A Design Principle for a Post-translational Biochemical Oscillator”.Cell Reports,2012,doi:10.1016/j.celrep.2012.09.006
発表者
理化学研究所
生命システム研究センター 細胞デザインコア 合成生物学研究グループ
グループディレクター 上田 泰己 (うえだ ひろき)
特別研究員 大出 晃士 (おおで こうじ)
お問い合わせ先
生命システム研究センター
広報担当 川野 武弘 (かわの たけひろ)
Tel: 070-6800-3938 / Fax: 06-6155-0112
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.自律振動子
外部から周期的な信号やエネルギーの供給がなくても、時間が進むにつれて周期的な変化を作ることができるシステム。例えば多くの電池式腕時計では、電池から送られるエネルギーを使い振動子(水晶発振子)から一定周波数の振動を生み出し時間を測っている。今回のタンパク質修飾に基づく自律振動子の場合は「基質の集団の修飾状態の時間周期的な変化を起こすシステム全体」を指す。 - 2.タンパク質の可逆的修飾
タンパク質を構成するアミノ酸に、リン酸基やメチル基などの分子が修飾酵素の働きによって付加されることをタンパク質の修飾と呼ぶ。また、多くの場合修飾は可逆的に制御されており、脱修飾酵素によって修飾が外される。これをタンパク質の可逆的修飾と呼ぶ。とりわけ、タンパク質の可逆的リン酸化(リン酸化と脱リン酸化)は、真核生物で広く用いられているタンパク質の機能調節機構である。 - 3.ミカエリス・メンテン式
ドイツのレオノール・ミカエリスとカナダのモード・メンテンによって1913年に提唱された酵素反応の速度を記述する式。今日まで多くの酵素反応がこの機構を用いて解析・解釈されてきている。 - 4.転写ネットワーク
遺伝子はDNAからmRNAに転写され、mRNAからタンパク質に翻訳されて機能する。翻訳されたタンパク質が自己もしくは他の遺伝子の転写翻訳を制御する場合、それらは相互に関係し合うネットワークを形成する。 - 5.転写因子の負のフィードバック制御
翻訳されたタンパク質が転写因子として機能し、自分の遺伝子の転写を抑制するように働く場合の制御機構。転写因子をAとすると、Aが転写される→Aの量が増える→Aの転写が抑制される→Aの量が減る→Aの転写抑制が弱まる→Aが再び転写される…という一連の流れにより、Aの量が増減を繰り返す振動が生み出される場合がある。 - 6.シアノバクテリアの概日時計
多くの生物の概日時計は、転写因子の負のフィードバック制御が振動子を形成していると考えられているが、シアノバクテリアでは転写を介さず、タンパク質のリン酸化と脱リン酸化で概日時計振動子が構成される。2005年に名古屋大学の近藤孝男教授のグループによって、シアノバクテリア概日時計に関わる3種類のタンパク質KaiA, KaiB, KaiCをATPとともに混合すると、KaiCのリン酸化状態がおよそ24時間周期で振動することが示された。KaiCタンパク質は自己リン酸化活性、自己脱リン酸化活性の双方を有する。すなわち、修飾酵素、脱修飾酵素、基質のすべての役割を一種類のタンパク質KaiCが果たしている。一般的に、これら3つの役割はそれぞれ異なるタンパク質が担うが、タンパク質の可逆的修飾だけで駆動される振動子はKaiC以外には見つかっていない。
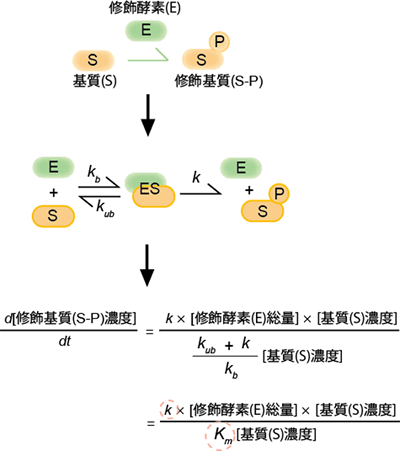
図1 ミカエリス-メンテン式とその反応機構の模式図
修飾酵素Eが、基質(S)を修飾基質(S-P)に変換する酵素反応を触媒すると想定する。ここでは、細胞内でよくみられるリン酸化を想定して修飾をP(リン酸:Phosphate)で表している。このプロセスでは、基質と修飾酵素が1:1で結合し、酵素基質複合体(ES)を作り、その後修飾基質が生成すると考える。基質酵素の結合定数をkb、基質酵素の解離定数をkub、基質酵素複合体から基質が修飾される反応速度定数をkとする。基質酵素の結合と解離が、反応速度よりも十分に早いとき、修飾基質が作られる速さは、下段の式で近似される。kは酵素が修飾・脱修飾を受ける速度、Km=(kub+k)/kbは酵素-基質の親和性のそれぞれの指標となる定数である。
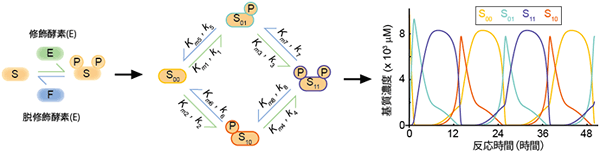
図2 基質の2カ所が修飾を受ける場合に生じる周期的な基質修飾状態の変化
基質の2カ所が修飾酵素と脱修飾酵素によって可逆的修飾を受けると想定する。この時、基質の修飾状態は4通りある(修飾を受けていないS00、片方のみ修飾されたS01とS10、両方修飾されたS11)。基質の修飾状態によって、修飾反応(緑線)と脱修飾反応(青線)は異なる酵素-基質親和性Kmと反応速度kで生じるとする。このモデルでは、各ステップのKmとk値の組み合わせによって、基質状態が時間とともに周期的に変化する場合がある。
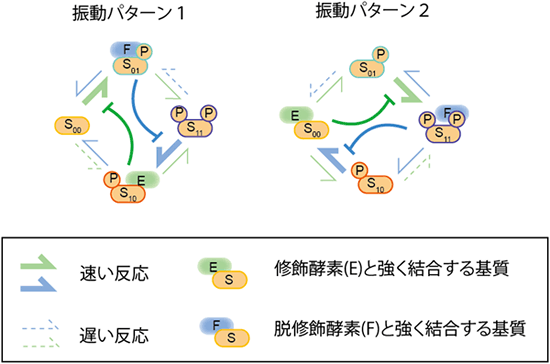
図3 修飾状態の振動を作る親和性と反応速度の特徴
振動を作る親和性と反応速度の組み合わせを12億通り解析することで見いだした2つの特徴。より速い傾向にあった反応速度を太線で、遅い傾向にあった反応速度を点線で表し、酵素と基質の親和性が高い状態には、それぞれ修飾・脱修飾酵素を表示した。1つの特徴として、対となる速度の速い修飾反応・脱修飾反応の方向の向きが揃っていることがあげられる。つまり右回りの反応速度が、左回りの反応速度よりも早い。2つ目の特徴として、特定の修飾状態の基質が酵素と高い親和性があることで、次の反応ステップを阻害する(T字マーク)ことが挙げられる。
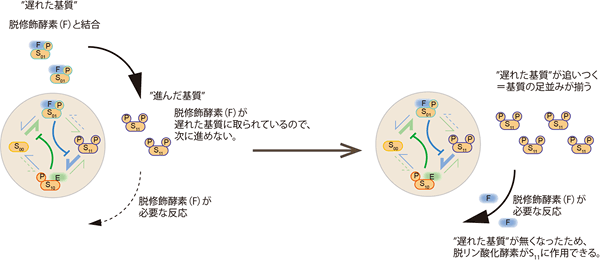
図4 基質集団の修飾状態の同調の模式図
振動が生じるためには、基質集団の修飾状態が時間ごとにそろう必要がある。今、基質修飾状態がS01とS11に分散しているとする。右回りの反応速度が高いために、S01は修飾されS11になる傾向にある。S11は脱修飾を受け、S10になる傾向が強いが、脱修飾酵素はS01と強く結合しており、S11の脱修飾反応を触媒することができない。従って、S01はS11に修飾されるが、S11は次の修飾状態(S10)に進むことができない。ほぼ全てのS01がS11に変換されたときに、脱修飾酵素はS01から解放され、S11を脱修飾する。こうして、S01とS11に分散されていた基質集団がS11にそろう。
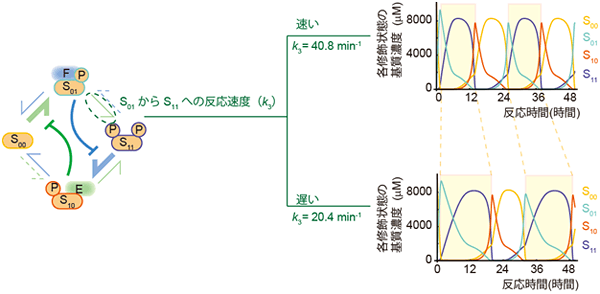
図5 酵素と強く結合する基質を消費する反応ステップが振動の周期決定に重要
S01からS11に至る反応速度k3を変化させた場合の振動の様子を示した。S11がS10へと進むためには脱修飾酵素FがS01から解離することが必要である。また、FがS01から解離するためにはS01がS11に変換されることによって減少する必要がある。k3はこのS01からS11への変換の速度定数であるため、k3が遅ければFは遅くS01から解離されるため、S11で停止している時間(黄色背景)が12時間から約20時間に長くなる。つまり、k3によって周期の長さ変化することが分かった。
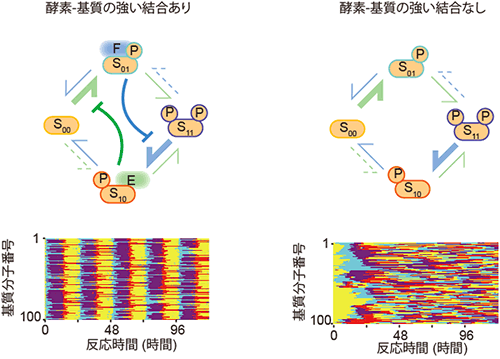
図6 酵素-基質の高い親和性を無くすと、基質集団の修飾状態が分散する
酵素-基質の親和性が高い場合と、低い場合それぞれについて修飾状態の変化を計算した。ここでは、基質分子の修飾状態の変遷を確率的に生じるものとして計算し、分子1つ1つを個別に計算している。親和性が高い場合(左)、それぞれの基質の修飾状態は各時間でよくそろっているが、低い場合(右)は修飾状態が分散している。