要旨
理化学研究所(理研)環境資源科学研究センター代謝システム研究チームの小山内崇研究員、平井優美チームリーダーらの研究グループ※は、微細藻類「ラン藻」を遺伝子改変し、代謝能力や光応答反応を変化させることに成功しました。これにより、アミノ酸の生産を効率的に増加できることが分かりました。
ラン藻は植物と似た光合成を行い、光エネルギーと二酸化炭素を使い、バイオプラスチックや糖、アミノ酸、色素などの有用物質を作ることができます。ラン藻の光合成メカニズムを分子レベルで理解できれば、将来的に光と二酸化炭素を使ったものづくりにつながると期待され、有用物質の効率的な生産方法の開発が求められています。ラン藻種に保存されている情報を伝達するタンパク質「ヒスチジンキナーゼ[1]」の1つであるHik8は、代謝酵素の発現を制御する可能性があるとされていましたが、代謝の仕組みに対し、実際にどのような影響を与えるかは分かっていませんでした。
研究グループは、淡水性ラン藻「シネコシスティス[2]」の遺伝子を改変し、細胞内のHik8タンパク質量を増加させたHik8過剰発現株を作製しました。このHik8過剰発現株の細胞内代謝産物を測定したところ、炭素の貯蔵源であるグリコーゲンが大きく減少することや、グリシン、アスパラギン酸など、光の条件に応じて7種類のアミノ酸が増加することが分かりました。次に、代謝変化の原因を調べるために、代謝の制御に重要なタンパク質「SigE[3]」の量を測定しました。その結果、Hik8過剰発現株では対照株(野生株)と異なり、暗条件下でもSigEが分解されないことが分かりました。つまり、Hik8タンパク質によって、暗条件下でもタンパク質や転写産物の分解が抑制されるなど、光応答性が変化することが明らかになりました。
今回の成果は、微細藻類の転写と代謝を改変する新しい方法といえます。今後、ラン藻の分子メカニズムの理解が深まることで、アミノ酸や糖などの効率的な生産につながると期待できます。
本研究は、JST戦略的創造研究推進事業個人型研究さきがけ(藻類・水圏微生物の機能解明と制御によるバイオエネルギー創成のための基盤技術の創出)の一環として行われ、欧州の科学雑誌『Environmental Microbiology』のオンライン版(11月18日付け)に掲載されました。
※研究グループ
理化学研究所 環境資源科学研究センター
代謝システム研究チーム
研究員 小山内 崇 (おさない たかし)
チームリーダー 平井 優実 (ひらい まさみ)
細胞生産研究チーム
上級研究員 白井 智量 (しらい ともかず)
チームリーダー 近藤 昭彦 (こんどう あきひこ)
筑波大学 植物代謝生理学研究室
教授 鈴木 石根 (すずき いわね)
背景
二酸化炭素を利用したものづくりは、地球温暖化対策のみならず、枯渇が懸念される化石燃料の代替資源を創出する手段としても非常に重要です。環境、資源問題が取り返しのつかない段階に達する前に、優れた環境技術を構築しておくことは、現代社会が担うべき責務です。
ラン藻は、光合成を行う細菌で二酸化炭素を固定できます。二酸化炭素の固定には、光合成で得られた光エネルギーを利用することから、ラン藻を用いて、二酸化炭素と光を共に利用する技術の開発に注目が集まっています。さらにラン藻は、光エネルギーと二酸化炭素を使い、アミノ酸、糖、バイオプラスチック、色素など、有用な物質を生産する能力があり、これらの生産能力を高める新技術の開発が求められています。
「シネコシスティス(Synechocystis sp. PCC 6803)」は、遺伝子改変が容易であり、増殖が比較的速いことなどから世界中で広く研究されているラン藻種です。シネコシスティスの細胞の代謝など、分子メカニズムを理解することで、基礎生物学の進展のみならず、有用物質生産などの応用展開が可能になると考えられています。ラン藻種に保存されているタンパク質「ヒスチジンキナーゼ」の1つであるHik8は、細胞内外の変化に伴って自己リン酸化されて、下流のタンパク質にシグナルを伝えていく「情報伝達タンパク質」です。これまでの研究で、Hik8がシネコシスティスの代謝に関与する可能性が示唆されていましたが、実際に炭素や窒素の代謝にどのような影響を与えるかは分かっていませんでした。
研究手法と成果
研究グループは、遺伝子改変によりシネコシスティスのHik8タンパク質を増加させたHik8過剰発現株を作製しました。明条件と暗条件に置いたHik8過剰発現株と対照株(野生株)に対し、メタボローム解析[4]を行って細胞内の代謝産物量を調べたところ、以下の3つのことが明らかになりました。Hik8過剰発現株は、①炭素の貯蔵源である多糖類のグリコーゲンが大きく減少すること、②明条件では6種類のアミノ酸(グリシン、イソロイシン、スレオニン、プロリン、アスパラギン酸、グルタミン酸)の量が対照株に比べて優位に増加すること、③暗条件ではアミノ酸のリジンが対照株に比べて優位に増加し、糖代謝上流の糖リン酸は減少することです(図1)。
次に、Hik8過剰発現による代謝の変化を明らかにするため、代謝の制御メカニズムについて調べました。これまでに研究グループは、シネコシスティスの糖代謝の制御因子を同定しています。その中で、SigEタンパク質は、糖の分解反応である「糖異化」を促進する因子であることを突き止めています注)。そこで、Hik8過剰発現株のSigEタンパク質の量を測定したところ、①明条件下では野生株よりも少ないこと、②明条件下から暗条件下に移動させた場合、野生株ではSigEタンパク質が分解されるが、Hik8過剰発現株では分解されないことが分かりました(図2)。
このように、Hik8は、SigEという代謝を制御する因子の働きを抑制していることが明らかになりました。さらに、メタボローム解析の他に免疫沈降法や遺伝学的解析を併用してHik8過剰発現株調べたところ、Hik8が、KaiC1という概日時計タンパク質[5]と協調して代謝を制御していることも明らかになりました(図3)。これらのことから、Hik8過剰発現によって、光応答反応が変化し、さらにさまざまなタンパク質との相互作用を介しながら、ラン藻の代謝を変化させることが明らかになりました。
注)Takashi Osanai et al. 2011 Genetic engineering of group 2 sigma factor SigE widely activates expressions of sugar catabolic genes in Synechocystis species PCC 6803.J.Biol. Chem. 286, 30962-30971.
今後の期待
今回、ラン藻において、情報を伝達するHik8タンパク質を使った新しい転写と代謝改変の方法を発見しました。数種類のアミノ酸が増加するという結果は、ラン藻の代謝改変を進展させることで、二酸化炭素を利用した有用物質の生産につながる可能性を示しています。今後、このHik8過剰発現株を詳細に解析することで、ラン藻細胞の分子メカニズムという基礎生物学と、有用物質生産という応用研究に重要な知見が得られるかもしれません。
原論文情報
- Takashi Osanai*, Tomokazu Shirai*, Hiroko Iijima, Ayuko Kuwahara, Iwane Suzuki, Akihiko Kondo, Masami Yokota Hirai, "Alteration of cyanobacterial sugar and amino acid metabolism by overexpression hik8, encoding a KaiC-associated histidine kinase", Environmental Microbiol, 2014, doi: 10.1111/1462-2920.12715
*Equally contributed
発表者
理化学研究所
環境資源科学研究センター 統合メタボロミクス研究グループ 代謝システム研究チーム
チームリーダー 平井 優美(ひらい まさみ)
研究員 小山内 崇(おさない たかし)
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.ヒスチジンキナーゼ
二成分制御系と呼ばれるグループのタンパク質で、細胞内外の変化を感知する。Hik8はヒスチジンキナーゼの1つで、自身をリン酸化することで、下流のレスポンスレギュレーターに情報を伝達する。 - 2.シネコシスティス
淡水性、単細胞性のラン藻。形状は球型で、直径が約1.5マイクロメートル。窒素固定を行わない。1996年にラン藻種の中で最初に全ゲノム配列が決定された。比較的増殖が速く、相同組換えによる遺伝子の改変が可能であるなどの利点を有することから、モデルラン藻として広く研究に使われている。 - 3.SigE
ラン藻の1種であるシネコシスティスが有する9つのRNAポリメラーゼシグマ因子の1つ。シグマ因子とは、RNAポリメラーゼ内でDNAに結合し、遺伝子の転写を開始する働きを持つタンパク質。研究グループは、SigEは糖代謝の制御因子であるほか、PHBの合成を促進することを発見した。
参考:2013年7月16日プレスリリース
「ラン藻が作るバイオプラスチックの増産に成功」 - 4.メタボローム解析
細胞内の代謝産物を網羅的に解析すること。従来は、目的の代謝産物を個別に定量していたが、細胞全体の代謝を把握するために、キャピラリー電気泳動マススペクトロメトリー、ガスクロマトグラフィーマススペクトロメトリー、液体クロマトグラフィーマススペクトロメトリーなどを用いて、可能な限り多くの代謝産物を定量する手法。 - 5.概日時計タンパク質
細胞内で、およそ24時間の周期を刻むために必要なタンパク質のこと。恒常的な条件で24時間の周期を維持し、温度の変化に応答しない。ラン藻では、KaiABCという3つのタンパク質が概日時計の本体である。KaiCのリン酸化状態は、KaiA、KaiBタンパク質が存在すると、試験管内で24時間周期を刻むことが知られている。
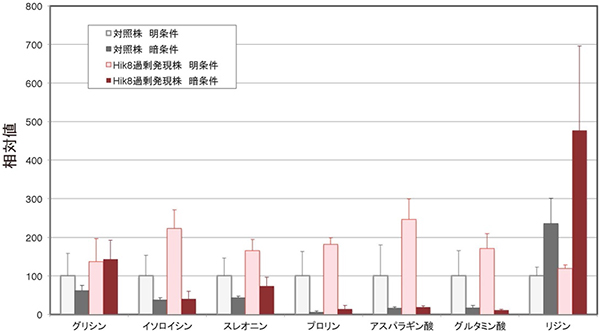
図1 メタボローム解析による7種類のアミノ酸量の比較
明条件または暗条件(1日)における対照株とHik8過剰発現株からそれぞれ抽出した7種類(グリシン、イソロイシン、スレオニン、プロリン、アスパラギン酸、グルタミン酸、リジン)のアミノ酸量の比較。対照株を100とした相対値で表している。どのアミノ酸もHik8過剰発現株で増加することが分かった。
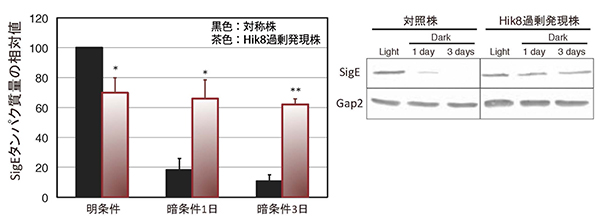
図2 SigEタンパク質量の比較
明条件、暗条件(1日または3日後)のSigEタンパク質量をウエスタンブロットという手法によって測定した。対照株を100とした相対値で表している。
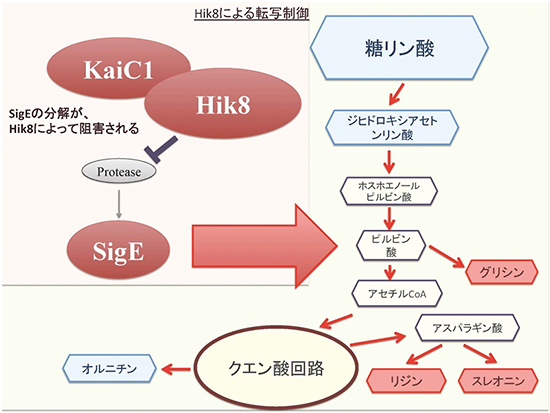
図3 暗条件でHik8過剰発現によって変化する代謝の模式図
メタボローム解析の結果より、Hik8は、概日時計タンパク質のKaiC1と協調して、糖の分解反応を促進していることが分かった。暗条件下で、Hik8過剰発現によって増える代謝産物を赤字、減る代謝産物を青字で示した。