要旨
理化学研究所(理研)小林脂質生物学研究室 小林俊秀主任研究員および上田善文客員研究員らの研究チームは、蛍光共鳴エネルギー移動(FRET)[1]を基にしたジアシルグリセロール(DAG)[2]蛍光プローブを用いることで、細胞内膜の裏側(外層:ルーメン側)のDAGを、表側(内層:細胞質側)のDAGとは独立かつリアルタイムに観察することに成功しました。
細胞膜をはじめとした脂質二重膜の表裏では、その脂質組成は非対称です。細胞は、この非対称性を巧妙に利用してアポトーシスや細胞の凝集などの細胞応答を制御します。機能性脂質分子の一種であり、異常な増加によって癌やアルツハイマー病などを引き起こすDAGは、細胞内膜の細胞質側とルーメン側に存在するため、両方の動態を明らかにすることが求められていました。しかし、従来の手法では、ルーメン側のDAGを観察することは不可能だったため、新たな測定手法の開発が求められていました。
研究チームは、ルーメン側に局在するp24タンパク質[3]の膜局在ドメインをプローブに連結した「Daglas-lum」というプローブを開発し、ルーメン側のDAGを観察することに成功しました。具体的には、細胞内でカルシウム濃度が上昇するとルーメン側でDAGが産生されることが分かりました。一方、同じ測定条件下では、研究チームが既に作製していた細胞質側のDAGを観察するためのプローブである「Daglas-em」には変化がありませんでした。これは、カルシウム濃度上昇によって、細胞内膜の細胞質側とルーメン側とで非対称なDAG産生が起きたことを意味しています。これは、生体がDAGの非対称な産生を利用して、さまざまな細胞応答を制御している可能性を示唆するものです。
本研究は、英国のオンライン科学雑誌『Scientific Reports』(8月12日号)に掲載されました。
※研究チーム
理化学研究所 小林脂質生物学研究室
主任研究員 小林 俊秀(こばやし としひで)
客員研究員 上田 善文(うえだ よしぶみ)
東京大学 理学系研究科化学専攻
名誉教授 梅澤 喜夫(うめざわ よしお)
東京大学 大学院総合文化研究科
准教授 佐藤 守俊(さとう もりとし)
金沢医科大学血液免疫内科学
教授 岡崎 俊朗(おかざき としろう)
特定講師 小木曽 英夫(おぎそ ひでお)
背景
細胞膜は、外層と内層の脂質二重層から成り立っており、その脂質組成は非対称です。例えば、糖脂質、スフィンゴミエリン、ホスファチジルコリンなどの脂質分子は外層に局在し、ホスファチジルエタノールアミンやホスファチジルセリンなどの脂質は内層に存在します(図1)。細胞は、この非対称性を巧妙に利用して、細胞応答を制御します。細胞がアポトーシスを起こす際には、内層にあったホスファチジルセリンが外層に露出し、eat-meシグナルとなることでマクロファージに認識され、貪食されます。また、血小板は、必要時にホスファチジルセリンやホスファチジルエタノールアミンを外層に露出させ、自己の凝集を誘導します。
ジアシルグリセロール(DAG)は、ホルモンや神経伝達物質に応答して産生する機能性脂質分子で、DAGの異常な産生は癌やアルツハイマー病を誘導することが知られています。DAGを産生する酵素は、細胞内膜(ミトコンドリア膜、小胞体膜、ゴルジ体膜)の内層(細胞質側)にも存在しますが、興味深いことに外層(ルーメン側)にも存在することが知られています。これは、DAGが細胞質側とルーメン側の両方の膜に存在することで、特別な機能を果たしている可能性があることを示唆しています。しかし、これまでルーメン側のDAGを観察する技術は存在しませんでした。
研究手法と成果
上田らは、2006年に細胞質側のDAGを観察するための蛍光共鳴エネルギー移動(FRET)を基にしたプローブである「Daglas-em」を開発しています(図2)注1)。このプローブの特徴は、結合する生体膜を変えることによって、興味のある膜でDAGを観察することができることです。
今回、ルーメン側のDAGを測定するために、ルーメン側に局在するp24タンパク質の膜局在ドメインをプローブに連結し、このプローブを「Daglas-lum」と名付けました。腎臓由来の培養細胞にDaglas-lumを発現させると、小胞体およびゴルジ体膜などに局在することがわかりました。つまり、Daglas-lumを用いて、小胞体およびゴルジ体のルーメン側のDAGを観察することができます。Daglas-lumと、細胞質側のDAGを観察するためのプローブであるDaglas-emを用いて細胞内膜の細胞質側とルーメン側のDAGの産生パターンを観測しました。その結果、細胞の機能に重要な役割を果たすカルシウムの細胞内濃度上昇に応じて、ルーメン側のみでDAGが産生されることが分かりました(図3)。さらに、ルーメン側のみでDAGが産生するメカニズムを調べるために、DAGを産生するさまざまな酵素の阻害剤を用いて調べた結果、このDAG産生は、ルーメン側に存在するスフィンゴミエリン合成酵素によって引き起こされていることが示唆されました。この結果は、細胞内膜の細胞質側とルーメン側においてDAGの産生は非対称的であることを示しています。
注1)Imaging diacylglycerol dynamics at organelle membranes. Sato M, Ueda Y, Umezawa Y.Nat Methods. 2006 Oct;3(10):797-9. PMID:16990811
今後の期待
研究チームは、細胞内膜のルーメン側のDAGを観察する技術を開発し、カルシウム濃度上昇によって細胞内膜のルーメン側のみでDAGが産生することを明らかにしました。これは、細胞内膜の細胞質側とルーメン側において非対称なDAG産生が起きたことを意味しており、生体がDAGの非対称な産生を利用して、さまざまな細胞応答を制御している可能性を示しています。
また、DAGの上昇は癌やアルツハイマー病に関与する可能性があり、今後、細胞質側だけでなくルーメン側のDAGの動態を詳細に調べることで、更なる病気の仕組みの解明につながると期待できます。また、今回の結果から小胞体内やゴルジ体にDAGと結合するタンパク質が存在することも示唆されます。
原論文情報
- Yoshibumi Ueda, Hideo Ogiso, Moritoshi Sato, Yoshio Umezawa, Toshiro Okazaki, and Toshihide Kobayashi, "Asymmetrical diacylglycerol dynamics on the cytosolic and lumenal sides of a single endomembrane in living cells", Scientific Reports, doi: 10.1038/srep12960
発表者
理化学研究所
主任研究員研究室 小林脂質生物学研究室
主任研究員 小林 俊秀(こばやし としひで)
 上田 善文
上田 善文
 小林 俊秀
小林 俊秀
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.蛍光共鳴エネルギー移動(FRET)
2つの蛍光分子がごく近接して存在する場合、一つの蛍光分子(ドナー)からもう1つの蛍光分子(アクセプター)へエネルギーが移行する。これを蛍光共鳴エネルギー移動(FRET)と呼ぶ。FRETの効率は2つの蛍光分子のスペクトルの重なりの大きさ、距離と角度により左右される。この原理を用いて、FRETを測定する事によりタンパク質相互作用、生化学反応や細胞内シグナル伝達の可視化を実現した。FRETはFluorescence Resonance Energy Transferの略。 - 2.ジアシルグリセロール(DAG)
ホルモンや神経伝達物質に応答して産生する機能性脂質分子で、細胞の増殖、分化などを制御する。DAGの異常な産生は癌やアルツハイマー病を誘導することが知られている。DAGはdiacylglycerolの略。 - 3.p24タンパク質
細胞内で作られるタンパク質(積み荷タンパク質)は、小胞輸送経路と呼ばれる道筋に沿って細胞内の目的地に運ばれます。p24タンパク質は、積み荷タンパク質の受容体で、小胞体、ゴルジ体およびその間の小胞のルーメン側に存在し、小胞体からゴルジ体へ向かう輸送小胞を制御します。
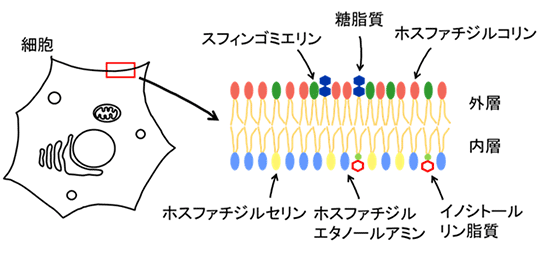
図1 細胞膜での脂質二重膜の外層と内層の脂質組成
脂質二重膜を構成する脂質は、外層と内層で異なっている。外層では、スフィンゴミエリン、糖脂質、ホスファチジルコリンが豊富に存在するのに対し、内層では、ホスファチジルセリン、ホスファチジルエタノールアミン、イノシトールリン脂質が存在する。
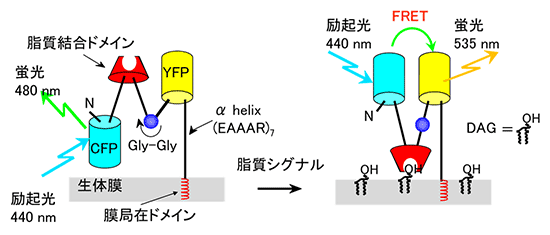
図2 蛍光共鳴エネルギー移動(FRET)DAGプローブ「Daglas-em」
青色蛍光タンパク質(CFP)、黄色蛍光タンパク質(YFP)をプローブ分子に導入する。DAGを選択的に認識する部位として、脂質結合ドメインを導入する。これらを剛直な、αヘリックス(黒線)で連結し、このヘリックスにGly-Glyを1箇所導入することで、ここを起点にプローブ分子が回転できるようにした。生体膜には、膜局在ドメインを用いて連結する。細胞膜内でDAGが増加すると、脂質結合ドメインはDAG、を捕捉し、CFP、YFP間の距離及び相対的配向の変化によってFRETが変化する。CFPは、440nmの波長で励起され480nmの蛍光を放出する。YFPは、FRETによって誘導された535nmの蛍光を放出する。この際の蛍光強度比変化(CFP/YFP)よりDAGの量を定量する。
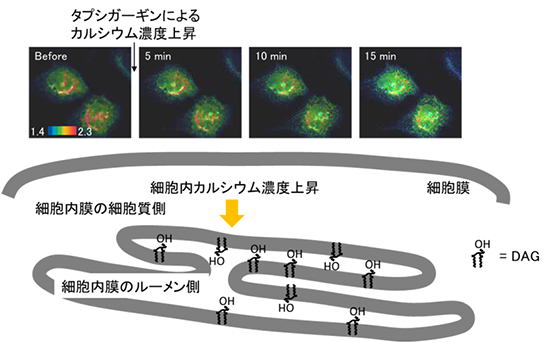
図3 細胞内カルシウム濃度上昇による細胞内膜ルーメン側のDAG産生
Daglas-lumを腎臓の培養細胞に発現させた。細胞質側からルーメン側にカルシウムを送り込む能力を妨げるタプシガーギンによって、細胞内のカルシウム濃度上昇を誘導すると、Daglas-lumの蛍光イメージが赤から青に変わり、DAGが産生したことを示した。細胞質側のDAGを測定するDaglas-emの場合は変化が観察されなかった。この結果からカルシウム濃度上昇によって細胞内膜のルーメン側でのみDAG産生が起きることが分かった。