2016年1月8日
理化学研究所
東京大学
オックスフォード大学
エジンバラ大学
植物Y染色体遺伝子地図を作成
-重イオンビームで作った変異体を使用、進化の過程でY染色体は逆位を起こしていた-
要旨
理化学研究所(理研)仁科加速器研究センター生物照射チームの阿部知子チームリーダー、風間裕介協力研究員、石井公太郎特別研究員と、東京大学大学院新領域創成科学研究科の河野重行教授らの共同研究グループ※は、重イオンビーム[1]で作り出した変異体と独自に開発したプログラムを用いて、ゲノム配列決定[2]が難しい植物Y染色体の遺伝子地図の作成に成功しました。
植物にはおしべとめしべを別々の個体につける「雌雄異株(しゆういしゅ)植物[3]」が存在し、ヒトと同様にXY型の性染色体を持つものがあります。植物の場合、両性花が祖先型のため、XY型でオスとメスを決めるには、XXでおしべを持たないメスであること、さらにY染色体にはおしべを伸ばしてめしべを引っ込めるという2つの性決定遺伝子が必要です。
ナデシコ科植物のヒロハノマンテマは、XY型の性染色体を持つ雌雄異株植物の代表例で、Y染色体には「めしべの発達を抑制する機能(GSF)領域」と「おしべの発達を促進する機能(SPF)領域」があり、これらの領域に性決定遺伝子が存在すると考えられています。しかし、ヒロハノマンテマのY染色体はヒトのY染色体の約10倍と巨大なため、ゲノム配列決定が困難で、性決定遺伝子の同定や性染色体の進化過程の解明は困難でした。そのため、ゲノム配列決定の基盤となるY染色体上の遺伝子の位置情報を記した遺伝子地図が求められていました。
共同研究グループは、ヒロハノマンテマに重イオンビームを照射し、Y染色体に変異を生じさせることで、おしべとめしべを両方持つ「両性花」や、どちらも持たない「無性花」といった変異体を多数作りました。これら変異体のY染色体上の欠失遺伝子を同定し、欠損遺伝子の一番もっともらしい並び順を推定する新規開発プログラム「DelMapper(デルマッパー)」を用いて、Y染色体の詳細な遺伝子地図の作成に成功し、GSF領域とSPF領域を識別することが可能になりました。
ヒロハノマンテマは、X染色体とY染色体に相同な遺伝子を持っています。以前に作成したX染色体の地図と比較したところ、Y染色体は進化の過程で遺伝子の並び順が全く逆になる巨大な逆位[4]を起こしていたことが分かりました。これは性決定遺伝子の進化を考える上で重要な知見です。
この成果は、ヒロハノマンテマY染色体の全ゲノム配列決定や性決定遺伝子の同定につながると期待できます。また、DelMapperは性染色体だけでなく、セントロメア[5](染色体の中央部分)付近など、遺伝学的な地図[6]の作製ができない染色体領域に存在する遺伝子の同定にも利用可能です。
本研究は、日本学術振興会科学研究費助成事業補助金基盤研究B「重イオンビーム欠失変異マッピングによる組換え抑制領域からの有用遺伝子同定」(代表者:風間裕介)、総合科学技術・イノベーション会議の戦略的イノベーション創造プログラム(SIP)「次世代農林水産業創造技術」(管理法人:農研機構 生物系特定産業技術研究支援センター、代表者:阿部知子)、および文部科学省科学研究費 新学術領域研究「生命科学系3分野支援活動、ゲノム支援」の一環として行われ、成果は英国のオンライン科学雑誌『Scientific Reports』(1月8日付け)に掲載されます。
※共同研究グループ
理化学研究所 仁科加速器研究センター 生物照射チーム
チームリーダー 阿部 知子(あべ ともこ)
協力研究員 風間 裕介(かざま ゆうすけ)
特別研究員 石井 公太郎(いしい こうたろう)
東京大学大学院 新領域創成科学研究科
教授 河野 重行(かわの しげゆき)
オックスフォード大学 植物科学部
准教授 ディミトリー・A・フィラトフ(Dmitry A Filatov)
エジンバラ大学 進化生物学研究所
名誉教授 デボラ・チャールズワース(Deborah Charlesworth)
背景
ナデシコ科植物のヒロハノマンテマはXY型の性染色体を持つ雌雄異株植物の代表例で、XYを持つとオス、XXを持つとメスになります(図1)。雌雄異株植物はおしべとめしべとを1つの個体につける両性花植物から進化したと言われています。XY型でオスとメスを決めるには、Y染色体上に少なくとも2つの性決定遺伝子が必要です。ヒロハノマンテマのY染色体には「めしべの発達を抑制する機能(GSF)領域」と「おしべの発達を促進する機能(SPF)領域」があり、これらの領域に性決定遺伝子が存在すると考えられています。しかし、ヒロハノマンテマのY染色体はヒトのY染色体の約10倍と巨大で、かつ同じ配列が反復した反復配列が多いため、ゲノム配列決定が進まず性決定遺伝子の同定や性染色体の進化過程の解明は困難でした。そのため、ゲノム配列決定の基盤となるY染色体上の遺伝子の位置情報を記した地図(遺伝子地図)が求められていました。
研究手法と成果
重イオンビームを植物や微生物に照射すると、効率よく突然変異体を作ることができます。上述のように、Y染色体はおしべやめしべなど花器官の形態に関与するため、花器官の形態が変わった変異体は、Y染色体に欠失が生じていると期待できます。そこで共同研究グループは、ヒロハノマンテマの花器官の変異体を複数種作りました。ヒロハノマンテマではY染色体のどこにあるか分からない遺伝子が数百個同定されています。各変異体でそれら遺伝子のうちどれが欠失しているかを調べ、欠失遺伝子の一番もっともらしい並び順を推定することができれば、Y染色体の遺伝子地図が描けると考えました。
共同研究グループは、理研の重イオン加速器施設「RIビームファクトリー(RIBF)[7]」で加速した重イオンビーム(炭素イオンビームやγ線)をヒロハノマンテマに照射し、Y染色体部分欠失変異体を40個体(①おしべとめしべを両方持つ両性花変異体15個体、②おしべもめしべも持たない無性花変異体9個体、③花粉が成熟しない変異体14個体、④めしべを持ちおしべを持たないメス様の変異体1個体、⑥Y染色体に欠失を持つオス1個体)を作り出しました(図2)。これら変異体に、自然突然変異で発生した無性花変異体1個体を加えた計41の変異体を解析に用いました。そして、各変異体について、Y染色体上の遺伝子69個のうちどの遺伝子が失われた(欠失変異を起こした)のかをPCR法[8]を使って調べました。
2つの遺伝子があった場合、互いの距離が近いほど1度の欠失で両方の遺伝子が失われる確率が高い、という先行研究注1)の考えを応用し、複数の遺伝子に欠失が見つかった場合、なるべくそれらの欠失が一ヶ所に集まるような並び順、すなわち、欠失の数がなるべく少なくなるような並び順を一番もっともらしいものと仮定しました。そして、そのような並び順を求めるために、巡回セールスマン問題[9]を応用した新プログラム「DelMapper(デルマッパー)」を開発し、Y染色体の遺伝子地図を作成しました(図3)。
遺伝子数、変異体数などの条件を変化させて作った仮想的な遺伝子地図を用意し、DelMapperに入力して正解の地図が描けるかどうかをテストすることで、作成した地図の正解率が90%と推定されました。作製した地図では、「めしべの発達を抑制する機能(GSF)領域」と「おしべの発達を促進する機能(SPF)領域」の位置をY染色体上にマップすることができました。さらに以前の研究で作成したX染色体の遺伝子地図注2)と比較したところ、Y染色体は進化の過程で大きな逆位を起こしていたことが分かりました(図4)。
注1)Cox et al. (1990) Radiation hybrid mapping: a somatic cell genetic method construction of high resolution maps of mammalian chromosomes. Science 250: 245–250.
注2)Kazama et al. SlWUS1; an X-linked gene having no homologous Y-linked copy in Silene latifolia. G3 (Bethesda) 2, 1269-1278
今後の期待
DelMapperは、性染色体だけでなく、セントロメア(染色体の中央部分)付近など遺伝学的な地図作成が困難な染色体領域に存在する遺伝子の同定にも利用できます。
本研究で得られた各遺伝子の位置情報は、Y染色体の全ゲノム配列を決定する上で役立ちます。ゲノム配列の決定により性決定遺伝子の同定が可能になると期待できます。
性染色体は常染色体から進化したと考えられています。ヒロハノマンテマのY染色体が誕生したのは1,000~2,000万年前であり、哺乳類のY染色体誕生が3億年前であるのと比べ極めて新しいといえます。本研究の成果は、性染色体の進化過程を調べる端緒となります。常染色体では、相同な染色体同士でDNA配列の交換(組換え)が起こりますが、逆位が生じると組換えができなくなります。共同研究グループが同定した逆位が、祖先Y染色体で生じ、X染色体との組換えが抑制されたことで、2つの性決定遺伝子をY染色体に定着させる引き金になった可能性が考えられます。
今後は、さらに詳細な地図を描くととともに、GSF遺伝子とSPF遺伝子を同定し、これら遺伝子が性を決定することの証明が重要です。そして、性決定遺伝子のそれぞれの進化速度[10]を調べることにより、どちらの遺伝子がどの順番で獲得され両性花から雌雄異株に進化したのかという、雌雄異株植物の進化の道筋が明らかになると期待できます。
原論文情報
- Yusuke Kazama, Kotaro Ishii, Wataru Aonuma, Tokihiro Ikeda, Hiroki Kawamoto, Ayako Koizumi, Dmitry A.Filatov, Margarita Chibalina, Roberta Bergero, Deborah Charlesworth, Tomoko Abe, Shigeyuki Kawano, "A new physical mapping approach refines the sex-determining gene positions on the Silene latifolia Y-chromosome", Scientific Reports, doi: 10.1038/srep18917
発表者
理化学研究所
仁科加速器研究センター 応用研究開発室 生物照射チーム
チームリーダー 阿部 知子(あべ ともこ)
協力研究員 風間 裕介(かざま ゆうすけ)
特別研究員 石井 公太郎(いしい こうたろう)
東京大学 新領域創成科学研究科
教授 河野 重行(かわの しげゆき)
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
東京大学大学院新領域創成科学研究科 総務係
Tel: 04-7136-5578 / Fax: 04-7136-4020
k-somu [at] adm.k.u-tokyo.ac.jp(※[at]は@に置き換えてください。)
補足説明
- 1.重イオンビーム
原子から電子をはぎ取って作られたイオンの中で、ヘリウムイオンより重いイオンを重イオンと呼ぶ。一方向に加速した重イオン粒子束を重イオンビームと呼ぶ。 - 2.ゲノム配列決定
シーケンサーで解読した約数百塩基対単位のDNA配列の情報を、パズルのようにつなぎ合わせて一続きのゲノム配列として決定する。一般に反復配列が多い領域では、多数のつなぎ方が存在する場合や間違ったつなぎ方をしてしまう場合があるため、配列の決定が難しい。 - 3.雌雄異株(しゆういしゅ)植物
雌雄が株ごとに完全に分かれている植物。オスはおしべを持ち、メスはめしべを持つ。 - 4.逆位
染色体上に生じる突然変異の1つ。染色体の一部分が反転してつなぎ変わったもの。 - 5.セントロメア
染色体のほぼ中央に位置する構造体。細胞分裂時には紡錘体がセントロメアに結合して、染色体を2細胞に分配する。 - 6.遺伝学的な地図
メンデルの独立の法則では、「個々の遺伝子は互いに干渉することなく、次の世代に分配される」とあるが、独立の法則に当てはまらない現象として「連鎖」が挙げられる。同じ染色体上にある遺伝子同士はまとまって遺伝するという現象である。しかし、連鎖している遺伝子でも、相同染色体同士が途中で交差して、遺伝子を入れ替えることで、連鎖を解消できる。これを組換えという。ある2つの遺伝子を考えた場合、「遺伝子間の距離が遠いほど、交差を起こすチャンスが増えるので組換えを起こす確率が高い」というこの原理を利用して、遺伝子間の距離を計算し、地図にしたのが「遺伝学的地図」である。ところが、組換え率は、染色体の場所によって異なる。セントロメア付近では組換えがほとんど起きないため、遺伝学的地図が描けない。 - 7.RIビームファクトリー(RIBF)
理研が有するRIビーム発生施設と独創的な基幹実験設備群で構成される重イオン加速器施設。生物に対する照射実験に用いるイオンは、炭素、窒素、ネオン、アルゴン、鉄の5種類。本研究では炭素イオンを照射した。 - 8.PCR法
ポリメラーゼ連鎖反応(polymerase chain reaction)の略。目的とするDNA断片(数百から数千塩基対)だけを選択的に増幅する技術。本研究では、Y染色体上の69個の遺伝子を増幅し、増幅されない遺伝子を欠失しているものとして判定した。 - 9.巡回セールスマン問題
組み合わせ最適化問題の1つ。任意の数の都市があり、それぞれの都市間の距離があらかじめ分かっているとする。セールスマンがある都市から出発し、残りの都市を全て訪問して出発した都市に戻るとき、移動距離が最小になるような都市の訪問順序を決定する問題を巡回セールスマン問題という。 - 10.進化速度
進化時間に対して塩基配列レベルで変化する速度。一定の期間内に遺伝子に蓄積される変異の数から求める。Y染色体には、変異が生じた場合に、正常な配列と交換して修復するための相同な染色体がないので、変異が生じやすい。Y染色体上の遺伝子の進化速度を比べることで、Y染色体上の遺伝子の新旧を調べることができる。

図1 ヒロハノマンテマのオスとメス
撮影:内田和歌奈(東京大学大学院新領域創成科学研究科博士課程3年)
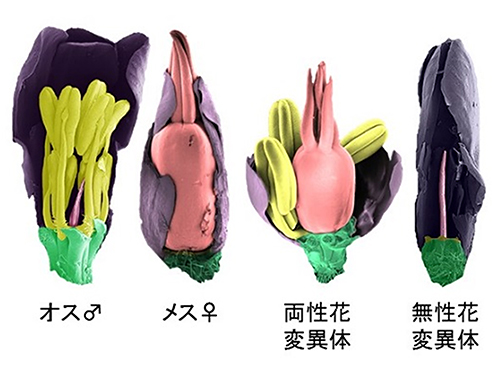
図2 ヒロハノマンテマの変異体の顕微鏡写真
野生型のオスはおしべを持ち、メスはめしべを持つ。重イオンビーム照射により、おしべとめしべとを両方持つ両性花変異体、どちらも持たない無性花変異体が得られた。両性花変異体は自家受粉して種をつけた。『Scientific Reports』より転載。
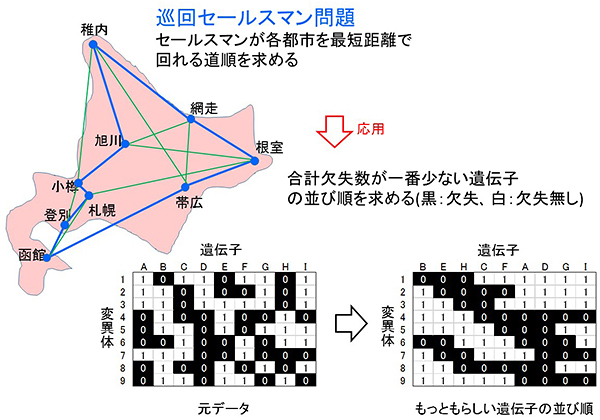
図3 巡回セールスマン問題を応用した新プログラム「DelMapper」で作成した遺伝子地図のイメージ図
セールスマンが複数の都市を1回だけ巡回する場合の最短経路を求める「巡回セールスマン問題」を遺伝子の並び方の決定に応用した新プログラム「DelMapper」の概要。元データでは、各変異体でどの遺伝子が欠失したか分かるが、並び順までは分からない。DelMapperを使用することで各変異体の合計欠失数が最小となる一番もっともらしい欠失遺伝子の並び順を求めることがきる。図に示した遺伝子地図のイメージ図では、B・E・H・C・A・D・G・Iが一番もっともらしい欠失遺伝子の並び順である。
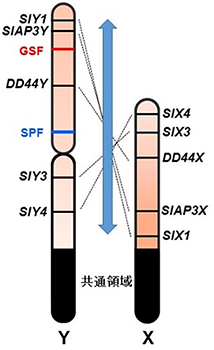
図4 Y染色体地図とX染色体地図の比較で示唆されたY染色体の逆位
以前に作成したX染色体の遺伝子地図と比較したところ、遺伝子の並び順がX染色体とY染色体とで全く逆になっていることが分かった。ヒロハノマンテマのY染色体は進化の過程でセントロメアを含む巨大な逆位を起こしたと考えられる。X染色体のセントロメアの位置は今のところ分かっていない。