2016年12月20日
理化学研究所
細胞死を司るカルシウム動態の制御機構を解明
-アービットが小胞体-ミトコンドリア間のCa2+の移動を制御-
要旨
理化学研究所(理研)脳科学総合研究センター発生神経生物研究チームの御子柴克彦チームリーダー、ベンジャミン・ボノー基礎科学特別研究員らの共同研究チーム※は、細胞内カルシウムチャネル[1]の制御因子「アービット(IRBIT)[2]」が、小胞体[3]からミトコンドリア[4]へのカルシウムイオン(Ca2+)の流入量をコントロールすることで、「アポトーシス」を制御することを発見しました。
アポトーシスはプログラムされた細胞死と呼ばれ、多細胞生物に見られる細胞の死に方の一つです。それは、不要になった細胞や損傷を受けた細胞が積極的に自滅することで、個体を健全な状態に保つために機能するメカニズムです。ストレス刺激などで細胞が損傷を受けると、エネルギー産生の場であるミトコンドリアへ過剰な量のCa2+が流入し、アポトーシスが誘導されます。このミトコンドリアへのCa2+流入は、細胞内のCa2+貯蔵庫である小胞体とミトコンドリアが接する場所である「小胞体-ミトコンドリア接触部位[5]」で起こります。このCa2+の動きには、小胞体の膜上にあるイノシトール三リン酸受容体(IP3R)[6]というカルシウムチャネルが重要な役割を果たしていますが、その制御メカニズムはよく分かっていませんでした。
今回、共同研究チームは、ゲノム編集技術[7]を利用してアービット(IRBIT)というタンパク質をコードする遺伝子を欠損したヒト細胞を作製・解析しました。その結果、アービット欠損細胞はアポトーシスが起こりにくいことが分かりました。また、小胞体-ミトコンドリア接触部位の構造に異常が見られ、小胞体からミトコンドリアへのCa2+の動きが阻害されていました。さらに「Bcl2l10[8]」というタンパク質がIP3RのCa2+放出活性を阻害してアポトーシスを抑制すること、アービットがBcl2l10の抗アポトーシス作用を抑制することを見いだしました。これらの結果は、アービットが小胞体-ミトコンドリア接触部位の形成、あるいは安定化を促進し、IP3Rを介したCa2+の動きを制御することにより、アポトーシスが誘導されることを示しています。
アポトーシスがうまく機能しないと、例えばDNAに損傷を受けてがん化した細胞が死なずに増殖してしまうなど、疾患の原因になります。今後、アービットとBcl2l10による細胞内Ca2+動態の制御機構をさらに解析することは、アポトーシスの機能不全よって引き起こされる疾患の分子機構の解明へつながると期待できます。
本研究成果は、英国の科学雑誌『eLife』に掲載されるのに先立ち、オンライン版(12月20日付け)に掲載されます。
本研究は、科学技術振興機構 戦略的創造研究推進事業 国際共同研究(ICORP)「カルシウム振動プロジェクト」(代表研究者:御子柴克彦)および発展研究(SORST)「カルシウム振動」(研究代表者:御子柴克彦)、日本学術振興会 科学研究費助成事業 基盤研究(S)の支援を受けて行われました。
※共同研究チーム
理化学研究所 脳科学総合研究センター 発生神経生物研究チーム
チームリーダー 御子柴 克彦(みこしば かつひこ)
基礎科学特別研究員 ベンジャミン・ボノー(Benjamin Bonneau)
研究員 安東 英明(あんどう ひであき)
研究員 河合 克宏(かわあい かつひろ)
テクニカルスタッフⅠ 廣瀬 松美(ひろせ まつみ)
北海道大学 医学系研究科 医学専攻 解剖学講座
准教授 岩永 ひろみ(いわなが ひろみ)
背景
「アポトーシス」はプログラムされた細胞死と呼ばれ、多細胞生物に見られる細胞の死に方の一つです。それは、不要になった細胞や損傷を受けた細胞が積極的に自滅することにより、個体を健全な状態に保つためのメカニズムです。アポトーシスがうまく機能しないと、本来は死滅すべき細胞、例えばDNAに損傷を受けてがん化した細胞が死なずに増殖してしまうなど、疾患の原因になります。
アポトーシスは細胞内のカルシウムイオン(Ca2+)によって制御されています。細胞が損傷を受けると、細胞内のカルシウム貯蔵庫となる細胞小器官である小胞体から、エネルギー生産の場であるミトコンドリアへCa2+が過剰に流入し、その結果アポトーシスが誘導されます。このミトコンドリアへのCa2+流入は、小胞体の膜とミトコンドリアの膜が接する場所(小胞体-ミトコンドリア接触部位)を介して行われます。小胞体からのCa2+の放出には、小胞体の膜上にあるイノシトール三リン酸受容体(IP3R)というタンパク質が重要な役割を果たしています。御子柴克彦チームリーダーらは、これまでにアービット(IRBIT)というタンパク質を発見し、アービットがIP3Rと結合し、その機能を制御していることを解明してきました注1,2,3)。
また、アポトーシスは、Bcl-2ファミリーというタンパク質群によっても制御されています。Bcl-2ファミリーには、アポトーシスを促進するグループと抑制するグループがあります。Bcl2l10というタンパク質はアポトーシスを抑制するグループに属すことは分かっていますが、詳しい機能は分かっていませんでした。
今回、共同研究チームは、アポトーシスの分子機構を明らかにするため、小胞体-ミトコンドリア間のCa2+の動きと、それを制御するアービットとBcl2l10の役割の解析を試みました。
注1)2006年6月13日プレスリリース「細胞内pHバランスの新たな制御機構の解明」
注2)2006年6月23日プレスリリース「細胞内のカルシウムチャネルに情報伝達を邪魔する“偽結合体”を発見」
注3)2014年4月14日プレスリリース「多動障害や社会行動の異常を抑える新しい分子機構を発見」
研究手法と成果
共同研究チームはまず、Bcl2l10がIP3Rに結合することによってIP3RのCa2+放出活性を抑制することを見いだしました。また、Bcl2l10はアービットとも結合し、アービットと協調してIP3RのCa2+放出活性を抑制することも分かりました。さらに、細胞分画法[9]やブルーネイティブゲル電気泳動法[10]で解析したところ、アービットとBcl2l10がIP3Rと共に、小胞体-ミトコンドリア接触部位に存在していることが分かりました。これらの結果から、通常、小胞体-ミトコンドリア接触部位のアービットとBcl2l10はIP3RのCa2+放出活性を抑制していると考えられます。
次に、ゲノム編集技術を用いて、アービットをコードする遺伝子を欠損したヒト細胞を作製しました。このアービット欠損細胞にアポトーシスを誘導するストレス刺激を加えたところ、野生型の細胞に比べて、アポトーシスが起こりにくくなることが分かりました。これにより、アービットはアポトーシスを促進する機能があると考えられます。
アービットはリン酸化(タンパク質の修飾の一種)されることにより、IP3Rと結合できる形になります。通常は、リン酸化型アービットはBcl2l10と共にIP3Rと結合し、IP3Rの機能を抑えています。ところが、細胞がストレス刺激を受けると、アービットが脱リン酸化されることが分かりました。脱リン酸化されたアービットは、IP3Rから解離します。その際に、アービットに結合しているBcl2l10も一緒にIP3Rから外れ、小胞体-ミトコンドリア接触部位から解離することが分かりました。アービットとBcl2l10による抑制が外れたIP3Rは、通常以上に多くのCa2+を放出します。その結果、小胞体からミトコンドリアに流入するCa2+量が増大し、ミトコンドリア内のCa2+濃度が上昇し、アポトーシスが誘導されると考えられます(図1)。
最後に、アービット欠損細胞の小胞体-ミトコンドリア接触部位を電子顕微鏡で観察したところ、野生型細胞に比べ、小胞体と接触しているミトコンドリアの割合が減少していることが分かりました。さらに、アービット欠損細胞では、小胞体からミトコンドリアへのCa2+の流入量が減少していました(図2)。この結果より、アービットは小胞体-ミトコンドリア接触部位の形成、あるいは安定化を制御する働きがあると考えられます(図1)。
これらの結果から次のモデルが考えられます。アービットは小胞体とミトコンドリアの接触を促し、小胞体からミトコンドリアへのCa2+の移動が効率よく行われるようにしています。一方で、アービットはBcl2l10と共にIP3Rの機能を抑制し、普段はCa2+の移動が過剰にならないように抑えています。ところが、細胞がストレス刺激を受けると、アービットは脱リン酸化されてBcl2l10と共にIP3Rから解離します。抑制が外れたIP3Rは、より多くのCa2+を放出します。その結果、ミトコンドリア内のCa2+濃度が上昇し、アポトーシスが誘導されます。これがアービットとBcl2l10がIP3Rを介してアポトーシスを制御する分子機構であると考えられます(図1)。
今後の期待
今回の研究により、アービットとBcl2l10がCa2+の動きをコントロールしてアポトーシスを制御していることが明らかになりました。がん細胞は、しばしば抗がん剤によるアポトーシスに耐性を示します。ある種のアポトーシス耐性を示すがん細胞株では、アービットの発現が低下していることが報告されています注4)。また、上皮性卵巣がん患者では、アービットの発現量が多いほど、予後がよいという報告もあります注5)。したがって、アービットの機能不全によりアポトーシスが正常に行われないことが、がんの発症あるいは増悪に関与している可能性があります。
今後は、アービットの脱リン酸化のメカニズムの解析、アービット欠損マウスを用いた解析など、アービットとBcl2l10による細胞内のCa2+動態の制御機構をさらに解析することで、アポトーシスの機能不全によって引き起こされるがんなどの疾患発症の分子機構の解明が期待できます。
注4)Wittig R, Nessling M, Will RD, Mollenhauer J, Salowsky R, Münstermann E, Schick M, Helmbach H, Gschwendt B,Korn B, Kioschis P, Lichter P, Schadendorf D, Poustka A. 2002. “Candidate genes for cross-resistance against DNA-damaging drugs.” Cancer Research62:6698–6705.
注5)Jeong W, Kim HS, Kim YB, Kim MA, Lim W, Kim J, Jang H-J, Suh DH, Kim K, Chung HH, Bazer FW, Song YS, Han, JY, Song G. 2012. “Paradoxical expression of AHCYL1 affecting ovarian carcinogenesis between chickens andwomen. Experimental” Biology and Medicine 237:758–767.
原論文情報
- Benjamin Bonneau, Hideaki Ando, Katsuhiro Kawaai, Matsumi Hirose, Hiromi Takahashi-Iwanaga, Katsuhiko Mikoshiba, "IRBIT controls apoptosis by interacting with the Bcl-2 homolog, Bcl2l10, and by promoting ER-mitochondria contact", eLife, doi: 10.7554/eLife.19896
発表者
理化学研究所
脳科学総合研究センター 発生神経生物研究チーム
チームリーダー 御子柴 克彦(みこしば かつひこ)
基礎科学特別研究員 ベンジャミン・ボノー(Benjamin Bonneau)
 発生神経生物研究チームのメンバー
発生神経生物研究チームのメンバー前列左から御子柴克彦チームリーダー、ベンジャミン・ボノー基礎科学特別研究員
後列左から廣瀬松美テクニカルスタッフⅠ、安東英明研究員、河合克宏研究員
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
科学技術振興機構 広報課
Tel: 03-5214-8404 / Fax: 03-5214-8432
jstkoho [at] jst.go.jp(※[at]は@に置き換えてください。)
JST事業に関すること
科学技術振興機構 研究プロジェクト推進部 大山 健志(おおやま たけし)Tel: 03-3512-3528 / Fax: 03-3222-2068
eratowww [at] jst.go.jp(※[at]は@に置き換えてください。)
補足説明
- 1.カルシウムチャネル
カルシウムイオン(Ca2+)は、真核細胞の最も普遍的な情報伝達物質の一つである。カルシウムチャネルはCa2+の通り道となるタンパク質の総称で、細胞内のCa2+濃度を制御している。 - 2.アービット(IRBIT)
御子柴克彦チームリーダーらが発見したタンパク質。イノシトール三リン酸受容体(IP3R)に結合し、その機能を制御するなど、多様な機能を持つ。IRBITは、IP3 receptor binding protein released with inositol 1,4,5-trisphosphateの略。 - 3.小胞体
細胞小器官の一つで、タンパク質の合成や品質管理、およびカルシウムの貯蔵庫として働く。 - 4.ミトコンドリア
細胞小器官の一つで、エネルギー産生を行う。 - 5.小胞体-ミトコンドリア接触部位
小胞体とミトコンドリアの膜が接触する場所で、小胞体膜上のタンパク質、ミトコンドリア膜上のタンパク質、それらをつなぐタンパク質などにより形成される。小胞体からミトコンドリアへ、エネルギー産生に必要なCa2+の移動が行われる。このCa2+の移動が過剰になると、アポトーシスが誘導される。 - 6.イノシトール三リン酸受容体(IP3R)
カルシウムチャネルの一つ。Ca2+の貯蔵庫である小胞体からCa2+の放出を行う。 - 7.ゲノム編集技術
配列特異的なDNA切断酵素を用いて、特定の遺伝子を破壊したり、改変したりする技術。2012年に報告されたCRISPR/Cas9を用いたシステムは、その簡便性と高効率性により急速に利用されるようになった。 - 8.Bcl2l10
アポトーシスを制御するBcl-2ファミリーに属するタンパク質。アポトーシスを抑制する働きがある。ビーシーエルツーエルテンと読む。 - 9.細胞分画法
細胞を破壊することにより、細胞小器官などの細胞内のそれぞれの構成要素を分離する方法。 - 10.ブルーネイティブゲル電気泳動法
タンパク質やDNAのような分子・粒子が電場中で移動することを利用して分離する方法を電気泳動法という。ブルーネイティブゲル電気泳動法は、タンパク質複合体を分離するための電気泳動法である。さらにSDSポリアクリルアミド電気泳動法と組み合わせることにより、タンパク質複合体を構成する個々のタンパク質を調べることができる。
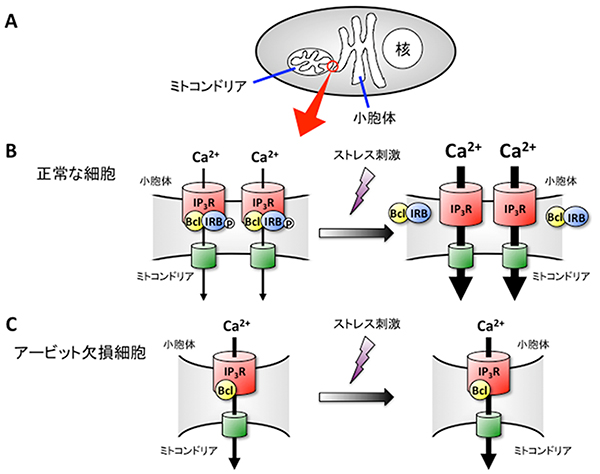
図1 アービットが小胞体-ミトコンドリア間のCa2+の動きを制御するメカニズム
A:細胞内の小胞体-ミトコンドリア接触部位を赤丸で示す。
B:正常な細胞の小胞体-ミトコンドリア接触部位の拡大図。(左)アービット(IRB)は、小胞体-ミトコンドリア接触部位の形成あるいは安定化を促進していると考えられる。また、リン酸化(P)されたアービットは、Bcl2l10(Bcl)と共にイノシトール三リン酸受容体(IP3R)に結合し、その活性を抑制している。小胞体からミトコンドリアへの適度なCa2+の移動は、ミトコンドリアの機能維持に必須である。(右)細胞がストレス刺激を受けると、アービットは脱リン酸化され、Bcl2l10を引き連れてIP3Rから解離する。抑制が外れたIP3Rは、過剰なCa2+を放出する。その結果、ミトコンドリア内のCa2+濃度が上昇し、アポトーシスが誘導される。
C:アービット欠損細胞の小胞体-ミトコンドリア接触部位の拡大図。(左)アービット欠損細胞では、野生型細胞に比べ小胞体と接触しているミトコンドリアの割合が減少し、小胞体-ミトコンドリア接触部位の幅が小さくなっている。そのため、ミトコンドリアへのCa2+の流入が効率的に行われない。(右)細胞がストレス刺激を受けても、Bcl2l10はIP3Rから解離しない。Bcl2l10はIP3R活性を抑制するため、ミトコンドリアへの過剰なCa2+流入が起こらず、アポトーシスは誘導されにくい。
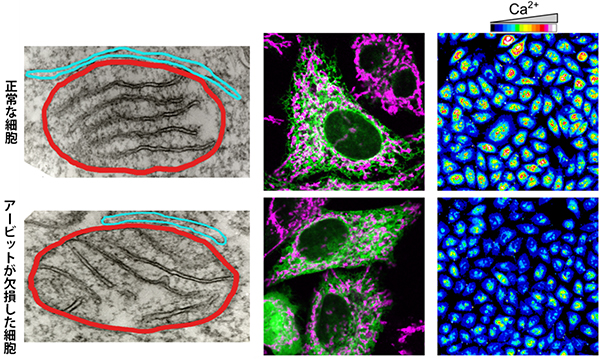
図2 正常な細胞とアービット欠損細胞の小胞体-ミトコンドリア接触部位とCa2+濃度の比較
左:電子顕微鏡像の比較。赤枠内がミトコンドリア、水色枠内が小胞体を示す。正常な細胞に比べ、アービットが欠損した細胞では、小胞体-ミトコンドリア接触部位(赤枠と水色枠の接触面)が減少していることが分かる。
中央:染色像の比較。紫がミトコンドリア、緑は小胞体を示す。この画像でも、正常な細胞に比べ、アービットが欠損した細胞では、小胞体-ミトコンドリア接触部位(紫と緑が重なる白い部分)が減少していることが分かる。
右:ストレス刺激後のミトコンドリア内のCa2+濃度の比較。正常な細胞に比べ、アービットが欠損した細胞ではCa2+濃度上昇が小さい。