2017年1月17日
理化学研究所
カリフォルニア大学アーバイン校
ウォーリック大学
新しいカルボキシル化酵素の発見
-天然有機化合物の構造多様性を創出する新規生合成経路の解明-
要旨
理化学研究所(理研)環境資源科学研究センター天然物生合成研究ユニットの高橋俊二ユニットリーダー、ケミカルバイオロジー研究グループの宮澤岳大学院生リサーチアソシエート(研究当時)、長田裕之グループディレクター、カリフォルニア大学アーバイン校のシェリル・サイ教授、ウォーリック大学のグレゴリー・チャリス教授らの国際共同研究グループは、天然有機化合物の構造多様性を作り出す新しい生合成[1]経路を発見しました。
微生物が生産する天然有機化合物は複雑な構造を持っています。それらの中から多くの抗菌薬、抗がん剤、抗寄生虫薬、免疫抑制剤などが発見されています。例えばポリケチド化合物[2]は、これら有用化合物の骨格を形成している化合物の一群でありポリケチド合成酵素(PKS)がその反応を触媒します。マロニルコエンザイムA(CoA)およびメチルマロニルCoAなどの主要伸長基質[2]に加えて、多様な側鎖を持つアルキルマロニルCoAを伸長基質に用いることで、生合成産物であるポリケチド化合物は多用な構造をとることができます。これまでにアルキルマロニルCoAは還元・カルボキシル化酵素(CCR)[3]によって生合成されることが知られています。しかし、ある放線菌[4]が生産するスタンボマイシン類はアルキルマロニルCoAをその骨格形成に用いているにも関わらず、その生合成遺伝子群の中にCCRに相同性を示す遺伝子が存在しません。国際共同研究グループは新しい生合成酵素の存在を予想し、その解明を試みました。
今回、国際共同研究グループは、ポリケチド化合物のスタンボマイシンおよびリベロマイシンの生合成機構を解析することによって、CCR反応を経由しないアルキルマロニルCoAの生合成経路を探索しました。その結果、反応中間体のアシルCoAをカルボキシル化[5]する「酵素SamR0483」が関与する新しい生合成経路を発見しました。また、生合成工学手法により、アルキンやアジド基といった細胞内での薬剤の追跡や標的タンパク質同定に役立つ“タグ”を持つ新しいスタンボマイシン類の創製に成功しました。
今後、本研究で見いだした生合成システムは化合物の構造多様化に寄与するだけでなく、天然有機化合物の作用機序解明に役立つ上述のタグを導入するツールとしての活用が期待できます。
本研究成果は、国際科学雑誌『Nature Communications』に(12月21日付け:日本時間12月22日)に掲載されました。
なお、本研究は日本学術振興会 科学研究費助成事業 基盤研究(B)の支援を受けて行われました。
背景
微生物が生産する天然有機化合物は複雑な構造を持っています。それらの中から、多くの抗菌薬、抗がん剤、抗寄生虫薬、免疫抑制剤などが発見されています。なかでもポリケチド化合物は、有用天然有機化合物の一群として知られており、抗菌薬(エリスロマイシン)、抗がん剤(ドキソルビシン)、抗寄生虫薬(エバーメクチン)は医薬品として利用されています。ポリケチド化合物の生合成はポリケチド合成酵素(PKS)がその反応を触媒し、生合成の開始基質に伸長基質を複数回つなぎ合わせることにより主骨格を形成します。PKSは複数の機能ドメイン(領域)で構成される巨大酵素であり、機能ドメインに応じて伸長基質が選択され、縮合、還元、脱水などの反応が起こります。これらの各機能ドメインを合成生物学的手法で組み立てることにより、自在に生理活性化合物を創出することが、ポリケチド生合成研究の究極の目標です。
骨格形成に用いられる伸長基質は、主にコエンザイムA(CoA)[6]にマロニル基を持つマロニルCoAとマロニル基にメチル基を持つメチルマロニルCoAですが、まれにアルキル側鎖に構造多様性を持つアルキルマロニルCoAが基質となる場合があります(図1)。そのためアルキルマロニルCoAは、ポリケチド化合物の構造多様化に寄与するツールとしての利用が期待されています。
これまでにアルキルマロニルCoAは、還元・カルボキシル化酵素(CCR)が(E)-2-エノイルCoAを還元・カルボキシル化することによって生合成されることが知られています。しかし、放線菌Streptomyces ambofaciens ATCC23877の生産するスタンボマイシン類はアルキルマロニルCoAをその骨格形成に用いているにも関わらず、その生合成遺伝子群の中に、CCRに相同性を示す遺伝子が存在していません。そこで、国際共同研究グループは新しい生合成酵素の存在を予想し、その解明を試みました。
研究手法と成果
高橋俊二ユニットリーダーらは、2011年にポリケチド化合物のリベロマイシンの生合成遺伝子クラスターを解明し注1)、その後、中鎖脂肪酸特異的アシルCoAリガーゼ[7](RevS)、還元・カルボキシル化酵素(RevT[3])が、アルキルマロニルCoA生合成に関与していることを明らかにしました注2)。スタンボマイシン生合成遺伝子群の中には、RevSに相同性を示すsamR0482遺伝子が存在しており、その近くには、カルボキシラーゼβサブユニットに相同性を示すsamR0483遺伝子が存在していました。そこで、SamR0482が中鎖脂肪酸をCoAにアシル基が結合したアシルCoAに変換後、SamR0483がカルボキシル化反応によりアルキルマロニルCoAを生合成すると予想しました(図2)。
遺伝子機能を知る上で重要な方法の一つに遺伝子破壊がありますが、スタンボマイシン生産菌ではsamR0483の遺伝子破壊が困難でした。そこで、生合成経路を解明するために、ヘプタン酸の3位を2個の重水素で標識[8]した(3-2H2)ヘプタン酸を合成しました。アルキルマロニルCoAがRevTに依存する従来の経路で生合成されるのであれば、アシルCoAデヒドロゲナーゼ反応(アシルCoA脱水素反応)により重水素が一つ欠落し、最終産物の分子量が1ダルトン(Da)[9]増加します。一方、RevTに依存しないカルボキシル化反応で生合成されるのであれば、アシルCoAデヒドロゲナーゼ反応を経由せず、最終産物の分子量が2 Da増加すると考えられます(図3)。
実際に、合成した(3-2H2)ヘプタン酸をStreptomyces ambofaciens ATCC23877の培養液に添加し、代謝産物を精製・解析した結果、スタンボマイシンの分子量が2 Da増加することを確認しました。このことから、スタンボマイシン生産菌ではアルキルマロニルCoAがカルボキシル化経路で生合成されることが示されました。
次に、SamR0483がアルキルマロニルCoAの生合成に直接関与しているかどうかを証明するため、異種放線菌にsamR0483遺伝子を導入する実験を行いました。放線菌Streptomyces sp. SN-593の生産するリベロマイシン類は多様なアルキルマロニルCoAを生合成しますが、これには、RevSとRevTが関与しています。また、revT遺伝子破壊株は、リベロマイシン類の骨格を形成するアルキルマロニルCoAが生合成できないため、リベロマイシン類が生産されません。SamR0483がアルキルマロニルCoAを生合成できるのであれば、revT遺伝子破壊株にsamR0483遺伝子を導入することによって、再びリベロマイシン類が生産されるようになると予想されます(図4)。
実際にsamR0483遺伝子をrevT遺伝子破壊株に導入したところ、リベロマイシン類の再生産が確認されました(図4)。さらに、(3-2H2)ヘプタン酸を上記のsamR0483遺伝子導入株に添加したところ、リベロマイシンD分子量の2 Da増加が確認されました。これらのことから、SamR0483はアルキルマロニルCoAの生合成に関与していることが明らかになりました。
さらに、SamR0483の基質認識機構を明らかにするため、SamR0483とヘキサノイルCoAの共結晶[10]を作製し構造解析を行いました。その結果、SamR0483はこれまでに報告されたプロピオニルCoAカルボキシラーゼβサブユニットと非常に類似した6量体構造をしていましたが、ヘキサノイルCoAのホスホパンテテイン(パントテン酸に由来するCoA構造の一部)および、アシル側鎖部分の認識に関わるアミノ酸が異なっていました。また、基質ポケット(基質が結合する部位)は疎水性の高いアミノ酸で構成され、基質入り口の柔軟性が高いことから、様々なアシルCoAを認識できる構造であることが明らかになりました。
また、SamR0483の共結晶構造解析の結果から、SamR0483は脂肪酸アシルCoA[11]を効率的にカルボキシル化することが示唆されたため、非天然型の脂肪酸を培養液に添加することにより、それらを取り込んだ新しいスタンボマイシン類の創製を試みました。その結果、アルキン(三重結合を一つ含む脂肪酸)である8-ノニン酸および、アジド基が結合した6−アジドヘキサン酸を取り込んだ新しいスタンボマイシン類の創製に成功しました。
注1)2011年プレスリリース「放線菌による「リベロマイシンA」生合成機序を遺伝子レベルで初めて解明」
注2)2015年プレスリリース「ポリケチド化合物の骨格形成に重要な酵素の機能解明」
今後の期待
本研究により、中鎖脂肪酸アシルCoAをカルボキシル化する新しい酵素によってアルキルマロニルCoAが生合成され、ポリケチド化合物のアルキル側鎖が形成されることを突き止めました。本研究で見出した生合成システムは、化合物の多様化に寄与するだけでなく、天然有機化合物の作用機序解明に役立つ“タグ”としてアルキンおよびアジド基などをポリケチド化合物に導入するツールとしての活用が期待できます。
原論文情報
- Lauren Ray, Timothy Valentic, Takeshi Miyazawa, David M. Withall, Lijiang Song, Jacob C. Milligan, Hiroyuki Osada, Shunji Takahashi, Shiou-Chuan Tsai & Gregory L. Challis, "A crotonyl-CoA reductase-carboxylase independent pathway for assembly of unusual alkylmalonyl-CoA polyketide synthase extender units", Nature communications, doi: 10.1038/NCOMMS13609
発表者
理化学研究所
環境資源科学研究センター 天然物生合成研究ユニット
ユニットリーダー 高橋 俊二(たかはし しゅんじ)
環境資源科学研究センター ケミカルバイオロジー研究グループ
大学院リサーチアソシエート(研究当時) 宮澤 岳(みやざわ たけし)
グループディレクター 長田 裕之(おさだ ひろゆき)
ウォーリック大学 化学科
教授 グレゴリー・チャリス(Gregory L. Challis)
カリフォルニア大学アーバイン校 分子生物学・生化学・化学・薬学科
教授 シェリル・サイ(Shiou-Chuan(Sheryl)Tsai)
 宮澤 岳
宮澤 岳
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
お問い合わせフォーム
産業利用に関するお問い合わせ
理化学研究所 産業連携本部 連携推進部
お問い合わせフォーム
補足説明
- 1.生合成
生物が作る酵素の作用によってさまざまな物質が合成されること。生物体を構成、維持する上で重要な化合物を一次代謝物、生育に必ずしも必須ではない化合物は二次代謝物と呼ぶ。 - 2.ポリケチド化合物、伸長基質
ポリケチド合成酵素(polyketide synthase, PKS)により、さまざまな開始基質にマロニルCoA[3]やメチルマロニルCoAなどの伸長基質が複数回縮合して生合成された天然有機化合物。PKSは複数の機能ドメイン(領域)を持つ巨大酵素であり、ドメイン構成に応じて還元、脱水等の反応が行われ構造が多様化される。 - 3.還元・カルボキシル化酵素(CCR)、RevT
還元反応とカルボキシル化反応を同時に触媒する酵素。クロトニルCoAをNADPH(ニコチンアミドアデニンジヌクレオチドリン酸)、CO2存在下で還元・カルボキシル化し、エチルマロニルCoAを合成する酵素(crotonyl-CoA reductase/carboxylase, CCR)として紅色細菌 Rhodobacter sphaeroidesから初めて同定された。その後、CCRはポリケチド化合物のアルキル側鎖を形成するための鍵酵素であることが明らかになっている。リベロマイシン生合成に関与する還元・カルボキシル化酵素としてRevTが知られる。 - 4.放線菌
土壌中など自然界に広く存在するグラム陽性の真正細菌であり、複雑な構造を持つ二次代謝産物を生産する。人類は、それらの中から、医薬、農薬、動物薬などの生理活性を持つ物質を利用してきた。医薬探索源として重要視されている。 - 5.カルボキシル化
有機化合物にカルボキシル基(-COOH)を導入する反応。 - 6.コエンザイムA(CoA)
酵素タンパク質と可逆的に結合することによって、酵素の活性を発現させる低分子量の有機化合物を補酵素(コエンザイム)という。コエンザイムAは生体内で重要な働きをする補酵素の一つである。 - 7.CoAリガーゼ
高エネルギー化合物であるATP(アデノシン三リン酸)を補酵素として、補酵素A(CoA)と基質のカルボキシル基を結合させる酵素。 - 8.重水素で標識
水素(1H、原子核は陽子1個のみ)の安定同位体である重水素(2HまたはD、原子核は陽子と中性子が1個ずつ)を用いて、生合成前駆体化合物を標識すること。これにより、二次代謝産物の生合成を予測することができる。 - 9.ダルトン(Da)
微小な質量を示す単位。静止して基底状態にある自由な炭素12(12C)原子の質量の12分の1と定義されている。 - 10.共結晶
タンパク質とそのタンパク質に結合する化合物を混合して、両者の複合体を結晶化すること。共結晶の構造解析をすることで、タンパク質と化合物の結合様式に関する情報を得ることができる。 - 11.脂肪酸アシルCoA
CoAのチオールが脂肪酸のカルボキシル基とチオエステル結合(-CO-S-)を形成した化合物。生体内において、脂肪酸代謝、エネルギー生産などに用いられる。
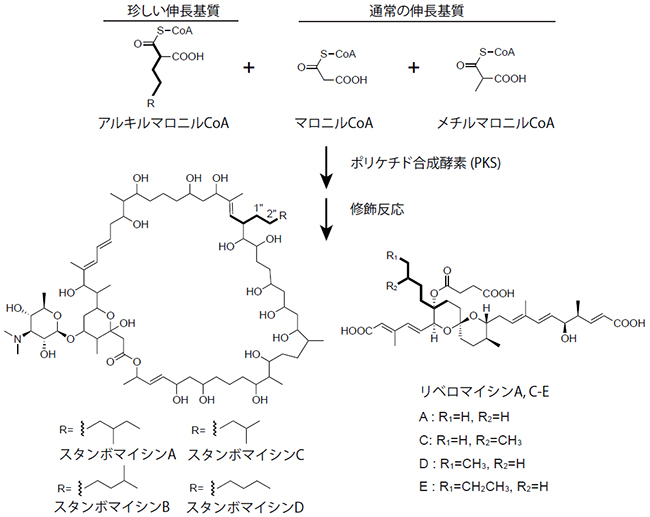
図1 ポリケチド化合物の生合成経路
ポリケチド化合物のスタンボマイシンA-D(左下)およびリベロマイシンA、C-E(右下)の生合成には、通常のマロニルCoAとメチルマロニルCoAに加え、珍しいアルキルマロニルCoAをポリケチド伸長基質として用いる。アルキルマロニルCoAの黒い太線(6-9炭素鎖)の骨格が、スタンボマイシンおよびリベロマイシンの太線部分に反映されている。
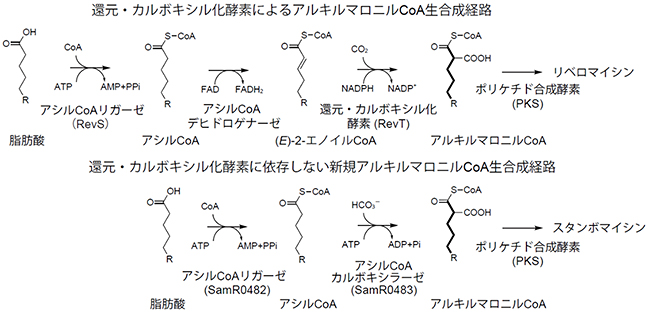
図2 アルキルマロニルCoAの2種類の生合成経路
ポリケチド化合物のリベロマイシン生合成では還元・カルボキシル化を経由するのに対し、スタンボマイシン生合成ではアシルCoAカルボキシラーゼ(SamR0483)により生合成されると予想した。
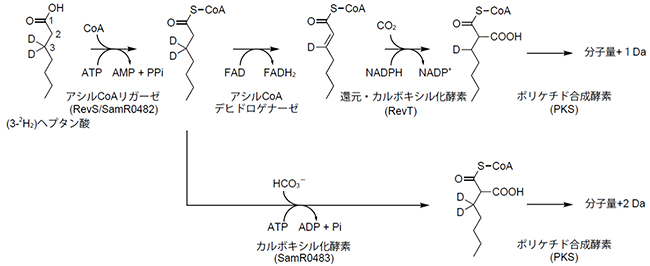
図3 (3-2H2)ヘプタン酸を用いたアルキルマロニルCoA生合成経路の解析
(3-2H2)ヘプタン酸を用いて、還元・カルボキシル化酵素(RevT)で生合成された場合、最終産物の分子量は1ダルトン(Da)増加するのに対し、カルボキシル化酵素(SamR0483)により生合成された場合、最終産物の分子量は2 Da増加する。なお、この反応式中では重水素2HはDで表記されている。
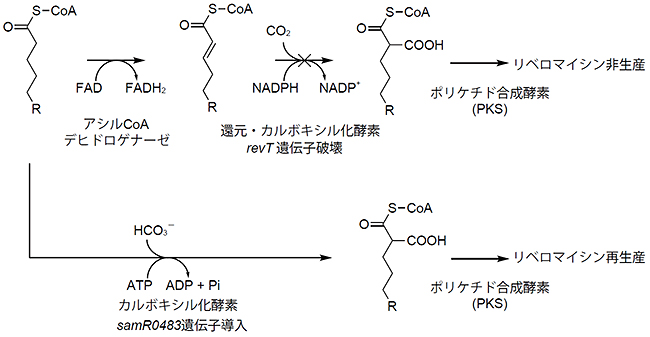
図4 SamR0483によるrevT 遺伝子破壊株の相補実験
revT遺伝子を破壊するとアルキルマロニルCoAが生合成できないため、リベロマイシン非生産となるが、samR0483遺伝子を導入することによりアルキルマロニルCoAが生合成され、リベロマイシンが再生産される。