2013年6月15日
独立行政法人理化学研究所
国立大学法人東京大学
植物の高い再生能力にはmRNAの制御機構が重要
-植物細胞の分化・増殖制御の理解から植物バイオテクノロジーの向上へ-
ポイント
- mRNA制御機構であるpre-mRNAスプライシングに関わる「RID1」を単離
- RID1の機能は植物特有の高い再生能力の基礎となる脱分化や器官再生に加えて発生に必須
- クローン増殖が難しい有用植物に有効なバイオテクノロジー開発への足がかり
要旨
理化学研究所(理研、野依良治理事長)と東京大学(濱田純一総長)は、植物の脱分化[1]・器官再生・発生において、遺伝子発現に必須な過程の1つであるmRNAからイントロン(遺伝子領域のうちタンパク質配列を指定しない領域)を除去する「pre-mRNAスプライシング[2]」と呼ばれるRNA制御機構が重要な働きを持つことを明らかにしました。これは、理研環境資源科学研究センター(篠崎一雄センター長)バイオマス工学連携部門 セルロース生産研究チームの大谷美沙都研究員、出村拓チームリーダーと東京大学大学院理学系研究科の杉山宗隆准教授による共同研究グループの成果です。
植物の一部を切り取って適切な条件で培養すると、植物細胞は比較的容易に脱分化してカルス(未分化の細胞塊)を形成し、再び根や茎に分化し完全な1つの植物体へと再生できます。こうした能力は、古くから農業や園芸においても組織培養や挿し木による優良苗のクローン[3]増殖に利用されています。しかし、どのような分子機構がこの性質を支えているのか、また植物の正常な発生制御とどう関連しているのかについては不明のままでした。
共同研究グループは、脱分化と器官再生が正常に起きないシロイヌナズナの変異体に着目し、その原因を詳細に調べました。その結果、RNAの絡みをほどく酵素RNAヘリカーゼ[4]の一種である「RID1」を新たに単離し、RID1が細胞核内にある核小体と呼ばれる領域に局在する新しいpre-mRNAスプライシング制御因子であること、RID1の機能が損なわれることで脱分化や器官再生が阻害されることを明らかにしました。さらにRID1の機能不全は、細胞分化や増殖を阻害するだけでなく、成長や発生に重要な分裂組織[5]確立、側根、配偶体形成などといった過程も阻害することも分かりました。これにより、植物に特徴的な高い再生能力にはpre-mRNAスプライシング制御が関わっていることが示されました。また、この制御能力が植物の特定な発生過程でとくに必要とされることが明らかになりました。
今回の研究は、組織培養の効率化につながる成果であり、改変ターゲット分子を明らかにしたことで、植物バイオテクノロジーの技術向上が期待できます。本研究成果は、米国の科学雑誌『The Plant Cell』に掲載されるに先立ち、オンライン版(6月14日付け:日本時間6月15日)に掲載されます。
背景
1つの細胞が体を形成するさまざまな種類の細胞全てに分化できる能力を持つことを分化全能性といいます。分化全能性を持つ細胞としては、動植物の受精卵細胞が挙げられます。植物細胞は、適切な条件で培養すれば再び根や茎に分化し完全な1つの植物体へと再生できるため、一般的にどの植物細胞も分化全能性を保持していると考えられています。これは、胚発生の段階で分化全能性を失う動物細胞と対称的であり、植物細胞の際立った特徴です。ただ、どのような分子機構によるものなのかについては、よく分かっていません。さらに、植物の正常な発生制御とどう関連しているのかという知見も限られています。
一方で、植物の高い再生能力は、古くから産業的に利用されてきました。とくに農業面では、挿し木や取り木、接ぎ木による優良苗のクローン増殖が利用されてきました。また園芸分野では、組織培養の一種である茎頂培養によって付加価値の高い花卉(かき)製品の増殖も行われています。しかし、植物種の中には容易にはクローン増殖できない種もあります。例えば、有用早生樹として利用されているユーカリやアカシアでは、挿し木時の発根効率が低いことが課題となっています。近年の環境問題の深刻化は、カーボンニュートラル[6]の理念に則った新たな産業形態の創出を求めていますが、そのためには有用植物の効率の良い増殖生産システムの構築が欠かせません。有用植物の分子育種の点からも、どのような分子メカニズムが再生能力と関連しているのかを明らかにすることが求められています。
そこで共同研究グループは、植物細胞が持つ高い再生能力を支えている分子機構の解明を目指しました。
研究手法と成果
共同研究グループは、モデル植物として広く研究されているシロイヌナズナを用いて温度感受性突然変異体[7]rid1-1(root initiation defective 1-1)を材料に分子遺伝学的解析を進めました。当初、この変異体は胚軸(発芽した幼植物体の茎部分)からの根の再生に関する変異体として単離されましたが、解析によってrid1-1変異体では胚軸脱分化(カルス形成)過程(図1)のほか、側根形成やシュート[8]再生の過程も高温になると阻害されることが分かりました。こうした表現型は、原因となる遺伝子であるRID1が、脱分化および器官再生において重要な役割を担っていることを意味しています(図2)。
RID1遺伝子について詳しく調べると、DEAH-box型RNAヘリカーゼと呼ばれるタンパク質をコードしていること、rid1-1変異体ではこの遺伝子配列のうちの1塩基が置換されることでRID1タンパク質を構成するアミノ酸の1つが別のアミノ酸に置換されていることが分かりました。このRNAヘリカーゼは、酵母やヒトで遺伝子からタンパク質が合成される過程で重要なpre-mRNAスプライシングに関連するタンパク質によく似ていました。そのため、RID1もpre-mRNAスプライシングに関与している可能性が考えられました。そこで、rid1-1変異体と野生型の細胞にタンパク質をコードしないイントロンを含む遺伝子を導入してスプライシング効率を比較しました。するとrid1-1変異体の細胞でスプライシング効率が著しく低下していることが分かりました(図3)。また、rid1-1変異体では選択的スプライシング[9]が起こることが知られている遺伝子の発現にも野生型とは異なるパターンがみられ、変異体ではmRNA上のどの領域をイントロンとして認識するかが乱れていることが分かりました(図4)。以上から、RID1はpre-mRNAスプライシング制御因子であることが示されました。興味深いことに、RID1は、pre-mRNAスプライシング進行の場である核質ではなく、核小体と呼ばれる核内小器官に局在しています(図5)。核小体はpre-mRNAスプライシングを担うsnRNPと呼ばれるRNA-タンパク質複合体形成の場であることから、RID1もsnRNP形成に関与していると考えられます。
また、共同研究グループは、RID1機能不全が植物発生にどういう影響をもたらすのかを詳細に解析しました。rid1-1は28°Cで育てると、芽生えの段階で致死となります。このとき、新たな器官を生み出す頂端分裂組織が正常に確立・維持されず、結果的に死に至っていることが分かりました。また、生存可能な22°Cで育てた場合でも、rid1-1では葉での維管束形成など細胞分化が異常になっていました。さらにRID1遺伝子の機能が完全に失われるノックアウト変異の解析を行ったところ、めしべで作られる卵細胞などの形成が異常となっていました。RID1遺伝子は、こうした変異体で異常が現れる器官や組織で強く発現していました。これらから、RID1が持つ機能、すなわちpre-mRNAスプライシング制御は、分裂組織形成や配偶体形成など、特定の植物発生制御で重要であることが示唆されました(図6)。
今後の期待
本研究によって、pre-mRNAスプライシング制御が、植物細胞に特徴的な高い再生能力を支える分子機構であることが明らかになりました。また、RID1機能は植物発生の過程にも必要とされることから、脱分化や器官再生などの二次的な細胞分化・増殖の過程と発生過程は、共通の分子基盤によることも分かりました。これは植物生理学的に重要な発見であると同時に、pre-mRNAスプライシング制御の生理機能を新たに突き止めたという点で、広く分子生物学的にも有意義な成果です。
共同研究グループはすでに、pre-mRNAスプライシング制御異常がなぜ細胞分化・増殖の阻害を引き起こすのか、その詳細なメカニズムの解明に取り組んでいます。その阻害の原因となっている分子が明らかになれば、植物細胞の分化や増殖を人為的にコントロールする技術開発につながると考えられます。これは、効率的なクローン増殖が難しい早生樹などの有用植物種に対して、有効な新規バイオテクノロジーとなることが期待できます。
原論文情報
- Ohtani, M., Demura, T., and Sugiyama, M. "ArabidopsisROOT INITIATION DEFECTIVE 1, a DEAH-box RNA helicase involved in pre-mRNA splicing, is essential for plant development." Plant Cell, 2013 doi: 10.1105/tpc.113.111922.
発表者
理化学研究所
環境資源科学研究センター バイオマス工学連携部門 セルロース生産研究チーム
チームリーダー 出村 拓 (でむら たく)
研究員 大谷 美沙都 (おおたに みさと)
お問い合わせ先
社会知創成事業 横断プログラム推進室
Tel: 048-462-1481 / Fax: 048-462-1220
報道担当
独立行政法人理化学研究所 広報室 報道担当
Tel:048-467-9272 / Fax:048-462-4715
東京大学大学院理学系研究科・理学部 広報室
広報室副室長(准教授) 横山 広美(よこやま ひろみ)
特任専門職員 武田 加奈子(たけだ かなこ)
Tel: 03-5841-8856 / Fax: 03-5842-1035
kouhou [at] adm.s.u-tokyo.ac.jp(※[at]は@に置き換えてください。)
補足説明
- 1.脱分化
生物の発生の過程で、細胞や組織が形態的・機能的に変化し、特定の機能が確立される過程を分化と呼ぶ。例えば受精卵が卵割を重ねて多細胞系となり、個々の細胞が特殊化した細胞(筋細胞や神経細胞など)などになることを指す。それに対して脱分化とは、既に分化した細胞が未分化の状態に変化すること。例えば植物のカルス(未分化の細胞の塊)形成などがある。 - 2.pre-mRNAスプライシング
遺伝子発現のためには、最初に遺伝子がタンパク質翻訳の鋳型であるmRNA(messenger RNA)へと転写される。転写直後のmRNAはpre-mRNA(プレmRNA)と呼ばれ、前駆体の状態。真核生物の場合、多くの遺伝子はエクソンとイントロンと呼ばれる2種類の領域を含んでいるが、このうちイントロンはタンパク質配列を指定しない領域で、最終的にはmRNAから取り除かれる必要がある。このイントロン除去過程をpre-mRNAスプライシングと呼ぶ。pre-mRNAスプライシングは核内で起こり、イントロンが除去された成熟mRNAは核外へ運ばれ、タンパク質に翻訳される。 - 3.クローン
同じ遺伝情報を持つ核酸や細胞、生物個体のこと。 - 4.RNAヘリカーゼ
絡み合った核酸をほどく働きをする酵素をヘリカーゼと呼び、DNAをほどくものはDNAヘリカーゼ、RNAをほどくものはRNAヘリカーゼと呼ばれる。最近では核酸-核酸間だけでなく、核酸-タンパク質間の相互作用を促進あるいは阻害する働きを持つものがあるなど、従来考えられてきたよりも幅広い機能を持つことが分かってきている。 - 5.分裂組織
陸上植物では細胞分裂は特定の組織で集中的に行われるが、この組織のことを分裂組織と呼ぶ。分裂組織の働きによって新たな細胞が供給され、さらに細胞が分化・伸長することで植物が成長する。 - 6.カーボンニュートラル
持続可能社会の構築に向けて提唱されている概念の1つで、二酸化炭素の排出と吸収の差がゼロになることをいう。具体的には、人間活動によって排出された二酸化炭素を植物などに吸収・固定させ、産まれたバイオマス由来資源による活動を行う、といった循環型システムを取ることで、大気中の二酸化炭素増減への影響を抑える構想。このためには、バイオマス資源の利活用性の向上が最重要課題となる。 - 7.温度感受性突然変異体
温度条件依存的に特徴的な表現型が観察される突然変異体のこと。高温で表現型が出る場合も、低温で表現型が出る場合もある。 - 8.シュート
茎とそれについている葉をまとめてシュート(shoot)と呼ぶ。シュートは茎の頂端に存在する分裂組織によって生み出される。 - 9.選択的スプライシング
タンパク質鋳型となるmRNAを作るときに、タンパク質配列指定に関わらないイントロンと呼ばれる領域を除去する機構がpre-mRNAスプライシングだが、1つのpre-mRNAから複数タイプの成熟mRNAが作られる場合を、とくに選択的スプライシング(alternative splicing)という。これによって、エクソン-イントロン構造などが変化したmRNAが生み出され、1つの遺伝子から複数のタンパク質が作られることもある。

図1 rid1-1変異体では温度依存的に胚軸からのカルス形成が阻害される
野生型(正常)およびrid1-1変異体の胚軸断片をカルス誘導培地上で22℃あるいは28℃で培養した。rid1-1では28℃で培養した場合にカルス形成が起こらない。
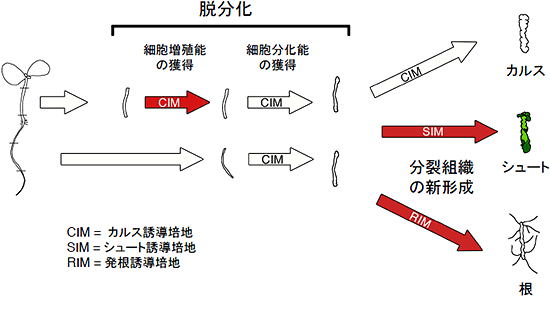
図2 脱分化・器官再生に関するrid1-1変異体の温度感受性のまとめ
rid1-1変異体で異常になる過程を赤矢印で示した。rid1-1変異体では胚軸からのカルス形成、シュート再生、および根の正常な発達が強く阻害される。こうした表現型から、RID1遺伝子は脱分化および器官再生に必要であると考えられた。
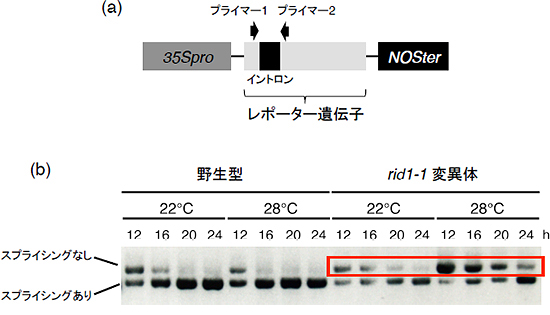
図3 rid1-1変異体では温度依存的にpre-mRNAスプライシング効率が低下する
- (a) 実験に用いた人工作製したレポーター遺伝子の模式図。人工的に遺伝子内にイントロンを挿入した。これを細胞内に導入することでpre-mRNAスプライシングが正常に起こると短い断片が、起こらないと長い断片がそれぞれ検出される。レポーター遺伝子とは、遺伝子組換えを行った時に導入した遺伝子が発現しているかどうか確認するために用いられる遺伝子のこと。
- (b) 実験の結果、野生型ではスプライシングが起こり短い断片が優勢となるが、rid1-1変異体ではスプライシングされていない長い断片がより多く検出された。また、この長い断片は28℃のときにより顕著に蓄積していた。
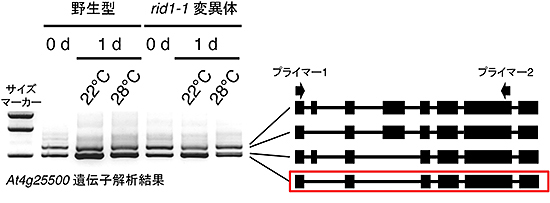
図4 rid1-1変異体では温度依存的に選択的スプライシングパターンが変化する
選択的スプライシングが起こることが知られているAt4g25500遺伝子について発現を調べたところ、rid1-1変異体では赤い四角で囲ったタイプのmRNAが減少していることが分かった。右図の黒い四角はエクソンとして認識された領域を、黒線はイントロンとして認識されて除去された領域を示す。

図5 RID1タンパク質は主に核小体に局在する
- (a) RID1タンパク質に蛍光タンパク質YFPを融合し、根の細胞で発現させた。緑色に見えるのがシグナル。
- (b) (a)と同じ細胞を蛍光染色試薬DAPIで染色した細胞像。DAPIは核酸と優先的に結合する試薬で、赤紫色に見えているのがシグナル。細胞中心の丸く見えている構造が核で、赤紫色が強い部分が核質、黒く抜けているように見える部分が核小体である。
- (c) (a)と(b)を重ね合わせた像。緑色のシグナルが核小体に存在しているのが分かる。
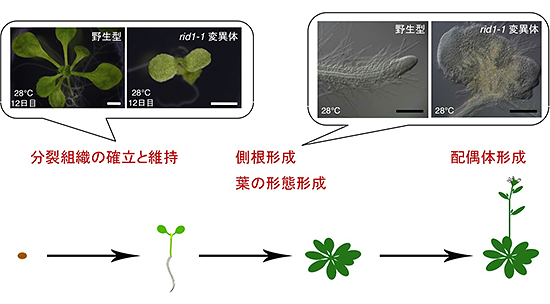
図6 RID1機能が必要とされる植物の発生過程のまとめ
rid1変異体で異常となる過程を赤字で示した。RID1機能はこれらの発生過程でとくに必要とされる。写真は28℃における野生型とrid1-1変異体の12日目の幼植物体の様子。野生型と異なり新しい葉が形成されない(左)、側根は形態が異常になり、こぶ状になる(右)。