要旨
理化学研究所(理研)統合生命医科学研究センター免疫転写制御研究グループの香城諭上級研究員、谷内一郎グループディレクターらの研究チームは、マウスを用いて、転写因子Bcl11bがヘルパーT細胞[1]、キラーT細胞[2]および制御性T細胞[3]への分化を適切に管理する重要な役割を担っていることを解明しました。
私たちの体を病原体やがんから守る際に重要な働きをするT細胞は、抗原を認識するT細胞受容体[4]の性質によって、ヘルパーT細胞、キラーT細胞、制御性T細胞などの細胞系列に分けられます。これらのT細胞は、骨髄に由来する共通の前駆細胞が胸腺[5]で分化・成熟することによって生まれます。それぞれの細胞に分化するためには、転写因子[6]として、ヘルパーT細胞ではThPOK、キラーT細胞ではRunx3、制御性T細胞ではFoxp3の発現が必要です。しかし、細胞系列に応じた転写因子の発現を適切に管理する機構についてはよく分かっていませんでした。
今回、研究チームは、ThPOKの発現を制御する転写因子を探索し、Bcl11bがその一つであることを突き止めました。通常ThPOKを発現した細胞ではRunx3の発現は抑制され、キラーT細胞への分化が抑制されます。しかし、Bcl11bを欠損したT細胞では、ThPOKを発現していてもRunx3の発現は抑制されず、ThPOKとRunx3を共に発現する異常なT細胞が出現しました。この異常な転写因子発現の結果、ヘルパー系列へ分化するはずの前駆細胞がキラーT細胞様細胞に、あるいはキラーT細胞へ分化するはずの前駆細胞がヘルパーT細胞様細胞に分化する異常が生じました。すなわち、ヘルパーT細胞とキラーT細胞の系列に応じた適切な転写因子の発現はBcl11bに管理されており、Bcl11bの欠損により転写因子が異常発現したことで、細胞の分化に混乱が生じたと考えられます。さらに、Bcl11bが制御性T細胞を分化誘導する転写因子Foxp3の発現にも関与していることも分かりました。このように、Bcl11bはさまざまな転写因子の発現を適切に制御することで、T細胞受容体の特徴に応じた分化を管理する役割を担う分子であることが明らかになりました。
近年、ES細胞(胚性幹細胞)[7]やiPS細胞(人工多能性幹細胞)[7]を用いた人為的なT細胞の作製による疾患治療の試みが進行しており、本成果は人為的T細胞誘導による免疫疾患の新たな治療法の開発に貢献するものと期待できます。
本研究は、国際科学雑誌『Nature Communications』(9月26日付け、日本時間9月26日)に掲載されます。
背景
私たちの体を、細菌やウイルスなどの病原体やがん細胞から守る細胞の一つにT細胞があります。T細胞には、免疫系の司令塔となるヘルパーT細胞、細菌・ウイルス感染細胞やがん細胞を排除するキラーT細胞、また過剰な免疫応答が生じないように免疫応答にブレーキをかける制御性T細胞などが存在します。各種T細胞は、骨髄に由来する共通の前駆細胞が、胸腺で行われるポジティブセレクション[8]という分化イベントによって、それぞれのT細胞が発現するT細胞受容体の性質に応じた細胞系列へ分化・成熟することで生まれます。
谷内グループディレクターらは、2002年にキラーT細胞への分化に重要な転写因子Runx注1)、2008年にヘルパーT細胞への分化に必要な転写因子ThPOK注2)に関する研究成果を報告しました。また、制御性T細胞の分化には、転写因子Foxp3の発現が必要であることも分かっています。これらの報告を通して、T細胞の運命決定にはそれぞれのT細胞系列に応じた転写因子の発現が必要であることが明らかになりました。しかし、それぞれのT細胞系列において、必要な転写因子の発現を適切に管理する機構の詳細についてはほとんど明らかになっていませんでした。
そこで研究チームは、ヘルパーT細胞の分化に必要なThPOKの発現を制御するタンパク質の同定を試みることから本研究を開始しました。
注1)Taniuchi I, et al. “Differential Requirements for Runx Proteins in CD4 Repression and Epigenetic Silencing during T Lymphocyte Development”Cell 111: 621-633, 2002
注2)2008年2月8日プレスリリース「ヘルパーかキラーか? Tリンパ球の分化運命決定のカギを発見」
研究手法と成果
谷内グループディレクターらは2008年の報告において、転写因子ThPOKの発現を制御する遺伝子領域を発見しています。研究チームはまず、この遺伝子領域に結合するタンパク質の同定を試み、転写因子Bcl11bが結合することを見いだしました。
次に、T細胞において特異的にBcl11bを欠損するマウスを作製・解析したところ、本来ThPOKを発現しない系列決定前の未熟なT細胞や成熟したキラーT細胞において、ThPOKが発現するようになり、ヘルパーT細胞に特異的なThPOKの発現は、Bcl11bによって制御されていることが分かりました。また、Bcl11bのC末端に存在するジンクフィンガー領域[9]を欠く変異マウスの作製によって、未熟なT細胞やキラーT細胞においてThPOKの発現が抑制されるためには、Bcl11bのC末端のジンクフィンガー領域が必要であることも分かりました。
通常、ThPOKを発現するヘルパーT細胞では、ThPOKの働きによってRunx3遺伝子の発現は抑えられます。しかし、Bcl11bを欠損するT細胞では、ThPOKを発現していてもRunx3の発現を抑制できなくなり、ThPOKとRunx3の両者を発現する異常な細胞が多数出現しました。
続いて、このように混乱した転写因子が発現する状態でのT細胞の分化状況を確認するために、キラーT細胞が存在しない主要組織適合抗原複合体(MHC)[10]クラスⅠ欠損マウスや、ヘルパーT細胞が存在しないMHCクラスⅡ欠損マウスとBcl11b欠損を組み合わせた解析を行いました。その結果、Bcl11bが欠損した状態では、MHCクラスⅠ欠損マウスにキラーT様細胞、またMHCクラスⅡ欠損マウスにヘルパーT様細胞が出現することが分かりました。すなわち、Bcl11bの機能不全によって、細胞の系列決定に必要な転写因子の発現に異常をきたし、その結果ヘルパーT細胞、キラーT細胞への細胞分化が混乱したものと考えられます(図1)。
次に、Bcl11bがRunx3遺伝子の発現をどのように制御しているのかを明らかにするために、クロマチン免疫沈降法と次世代シーケンサーを併用した解析法であるChIP-seq[11]という手法を用いて、Runx3遺伝子領域におけるBcl11b結合部位を確認しました。その結果、Runx3遺伝子プロモーター[12]の上流2カ所(-21kb、-39kb)にBcl11bの結合領域があることが分かりました。この領域がRunx3のエンハンサー[12]活性を持つこと、またこの領域にはRunx3を抑えるThPOKが結合することも分かりました。すなわち、ThPOKがこれらのRunx3エンハンサーに結合し、エンハンサー機能を抑制する段階において、Bcl11bが必要とされると考えられます。
さらに、Bcl11bを欠損するマウスを用いた解析によって、制御性T細胞の誘導に必要な転写因子Foxp3の発現においても、Bcl11bが重要であることを見いだしました。クロマチン免疫沈降を用いた解析の結果、Bcl11bはFoxp3の制御領域CNS3に結合し、Foxp3の発現誘導に重要な転写因子SATB1のCNS3への結合に関与していることが分かりました。
これらの結果から、Bcl11bが各系列に特異的な転写因子の発現を適切に管理する役割を担っていることが明らかになりました(図2)。また、Bcl11bの機能を喪失する時期を変化させたところ、T細胞の運命決定を担うポジティブセレクションの前の段階で機能を喪失させた場合に、より著しい転写因子発現異常がみられました。すなわち、Bcl11bはT細胞分化の早い段階でそれぞれの転写因子遺伝子領域に結合することによって、ポジティブセレクションシグナルを受け入れ適切な遺伝子発現へと転換するための準備を整える役割を担っていると考えられます。
今後の期待
今回、研究チームは転写因子Bcl11bが、T細胞の前駆細胞がヘルパーT細胞、キラーT細胞、および制御性T細胞への系列決定がなされる段階において重要な役割を担うことを見いだしました。系列特異的な転写因子の発現を管理する機構についてはこれまでほとんど明らかにされておらず、今回の発見はその解明の糸口になることが期待できます。
近年、がんなどの疾患を対象に、ES細胞やiPS細胞を用いた人為的なT細胞の作成による治療の試みが進行しています。T細胞分化を制御する機構の解明は人為的にT細胞を作製する方法の開発につながることから、本研究の成果は人為的T細胞誘導による免疫疾患の新たな治療法の開発に貢献するものと期待できます。
原論文情報
- Satoshi Kojo, Hirokazu Tanaka, Takaho A. Endo, Sawako Muroi, Ye Liu, Wooseok Seo, Mari Tenno, Kiyokazu Kakugawa, Yoshinori Naoe, Krutula Nair, Kazuyo Moro, Yoshinori Katsuragi, Akinori Kanai, Toshiya Inaba, Takeshi Egawa, Byrappa Venkatesh, Aki Minoda, Ryo Kominami and Ichiro Taniuchi, "Priming of lineage-specifying genes by Bcl11b is required for lineage choice in post-selection thymocytes", Nature Communications, doi: 10.1038/s41467-017-00768-1
発表者
理化学研究所
統合生命医科学研究センター 免疫転写制御研究グループ
グループディレクター 谷内 一郎(たにうち いちろう)
上級研究員 香城 諭(こうじょう さとし)
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
お問い合わせフォーム
産業利用に関するお問い合わせ
理化学研究所 産業連携本部 連携推進部
お問い合わせフォーム
補足説明
- 1.ヘルパーT細胞
免疫応答に関与するリンパ球、T細胞の一種。抗原の情報をB細胞へ伝えて抗体の産生を誘導したり、免疫応答を誘導する液性因子を放出したりすることにより、免疫反応の司令塔として働く。ThPOKという転写因子によって分化が制御されている。 - 2.キラーT細胞
免疫細胞の一種で細胞傷害性T細胞とも呼ばれる。ウイルス感染細胞やがん細胞など宿主にとって異物となる細胞を認識して破壊する。Runx3という転写因子によって分化が制御されている。 - 3.制御性T細胞
炎症やアレルギーの発端となる過度の免疫応答を抑制するT細胞亜集団。Foxp3という転写因子によって分化が制御されている。一般にT細胞は免疫活性化に働くが、制御性T細胞は他のT細胞の働きを抑制性に制御することにより、過剰なT細胞の働きを抑え、炎症の収束や自己免疫疾患発症抑制に重要な役割を果たす。 - 4.T細胞受容体
T細胞の表面に発現している受容体。主要組織適合抗原複合体(MHC)([10]参照)上に提示された抗原を認識する。α鎖とβ鎖の2本鎖で構成されており、それぞれのT細胞が一つの抗原に特異的なT細胞受容体を発現する。膨大なT細胞受容体の多様性は、T細胞受容体遺伝子の再構成によって形成される。 - 5.胸腺
T細胞が作られる臓器で心臓の上に位置している。骨髄から移動してきた前駆細胞が、胸腺の中でT細胞へと分化・成熟する。 - 6.転写因子
遺伝子の発現を調節するタンパク質。DNA上に存在する遺伝子の発現を制御する領域(プロモーター、エンハンサー、サイレンサーなど)に結合し、DNAを鋳型としてRNAが産生(転写)される時期や量を調節する。 - 7.ES細胞(胚性幹細胞)、iPS細胞(人工多能性幹細胞)
脊椎動物の初期胚が持つ、全ての種類の体細胞へ分化する能力を多能性という。多能性を持ち、試験管内で培養して無限に増やすことができる細胞を多能性幹細胞という。ES細胞は、哺乳類の着床前胚(胚盤胞)に存在する内部細胞塊から作製された多能性幹細胞である。iPS細胞は、成人の皮膚細胞などの体細胞・組織から採取した細胞に Oct3、 Sox2、 Klf4遺伝子などを導入して初期化し多能性を持たせ、人工的に作製した多能性幹細胞のこと。 - 8.ポジティブセレクション
胸腺において、機能的なT細胞を選別するイベントの一つ。主要組織適合抗原複合体(MHC)クラスⅠを認識するT細胞受容体を発現する細胞はキラーT細胞へ、MHCクラスⅡを認識するT細胞受容体を発現する細胞はヘルパーT細胞へ分化が誘導される。 - 9.ジンクフィンガー領域
発現を制御する遺伝子のプロモーター領域に、直接相互作用するタンパク質の領域のこと。四つのアミノ酸を介して亜鉛イオンと結合した分子構造で、DNAに相互作用する部分が「指」のような構造でDNAに結合するように見えることから、この名が付いた。 - 10.主要組織適合抗原複合体(MHC)
構成される分子から、クラスⅠとクラスⅡに分類される。T細胞受容体は、MHC上に提示される抗原を認識する。主として、MHCクラスⅠは細胞内の抗原(ウイルスや細菌成分など)、MHCクラスⅡは細胞外の抗原(外来の異物)をT細胞へ提示する。遺伝的な背景により、MHCには多型性があり、臓器移植の差異には、MHCが異なると拒絶反応が生じることから、主要組織適合抗原と呼ばれている。 - 11.ChIP-seq
ゲノムへのタンパク質結合を調べるクロマチン免疫沈降法と、次世代シーケンサーを組み合わせた網羅的解析方法。クロマチン免疫沈降で回収したDNA断片を、次世代シーケンサーを用いてゲノムワイドかつ網羅的に解析する。 - 12.プロモーター、エンハンサー
遺伝子の発現を調節するようなDNA領域。この領域に転写因子などのタンパク質が結合することで、遺伝子の発現が調節される。遺伝子の転写開始点から近い転写調節DNA領域がプロモーター、転写開始点からは隔たっている転写調節DNA領域がエンハンサーである。
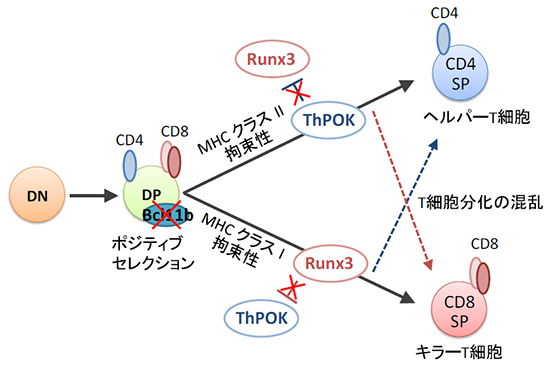
図1 Bcl11b欠損に伴う転写因子発現異常によって生じるT細胞分化の混乱
T細胞は胸腺内にて、細胞膜上にCD4/CD8分子を発現しないダブルネガティブ(DN)という未熟な段階を経てCD4/CD8を共に発現するダブルポジティブ(DP)へと成熟し、ポジティブセレクションを受ける。転写因子Bcl11bが存在する状態では、胸腺でのポジティブセレクションの後、主要組織適合抗原複合体(MHC)クラスⅡを認識するT細胞受容体を保有するT細胞は、ThPOKを発現しヘルパーT細胞へ、MHCクラスⅠを認識するT細胞は、Runx3を発現しキラーT細胞へ分化する(黒の矢印)。一方、Bcl11bを欠損すると、ヘルパーT細胞においてRunx3が、またキラーT細胞においてThPOKが発現され、T細胞分化に混乱をきたす(それぞれ青・赤の破線矢印)。
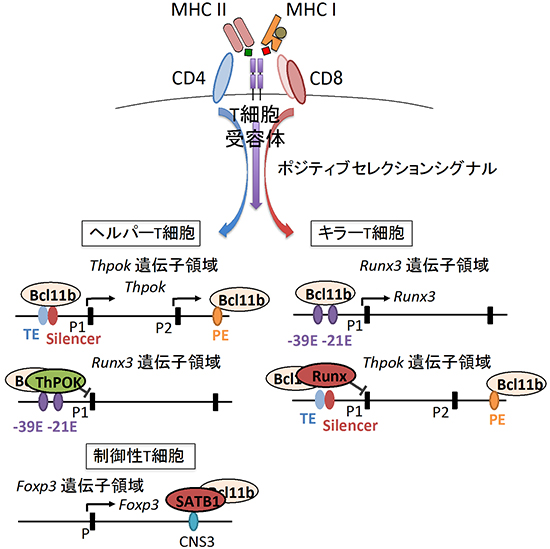
図2 Bcl11bによる転写因子発現制御機構
Bcl11bは、ThPOK遺伝子領域(TE、Silencer、PEなど)、Runx3遺伝子領域に存在する二つのエンハンサー領域(-39E、-21E)、Foxp3遺伝子領域に存在するCNS3領域に結合し、ポジティブセレクションシグナルを受け入れる準備を整えている。ポジティブセレクションによって入力されたT細胞受容体シグナルを、各系列に特異的な転写因子発現へと変換する役割を担うと考えられる。