Apr. 22, 2015 Press Release Biology
Look mom, no eardrums!
Studying evolution beyond the fossil record
Researchers at the RIKEN Evolutionary Morphology Laboratory and the University of Tokyo in Japan have determined that the eardrum evolved independently in mammals and diapsids—the taxonomic group that includes reptiles and birds. Published in Nature Communications, the work shows that the mammalian eardrum depends on lower jaw formation, while that of diapsids develops from the upper jaw. Significantly, the researchers used techniques borrowed from developmental biology to answer a question that has intrigued paleontologists for years.
The evolution of the eardrum and the middle ear is what has allowed mammals, reptiles, and birds to hear through the air. Their eardrums all look similar, are formed when the ear canal reaches the first pharyngeal pouch, and function similarly. However the fossil record shows that the middle ears in these two lineages are fundamentally different, with two of the bones that make up the mammalian middle ear—the hammer and the anvil—being homologous with parts of diapsid jawbones—the articular and quadrate. In both lineages, these bones connect at what is called the primary jaw joint.
Although scientists have suspected that the eardrum—and thus hearing—developed independently in mammals and diapsids, no hard evidence has been found in the fossil record because the eardrum is never fossilized. To overcome this difficulty, the research team and their collaborators turned to evolutionary developmental biology—or “evo-devo.” They noted that in mammals, the eardrum attaches to the tympanic ring—a bone derived from the lower jaw, but that in diapsids it attaches to the quadrate—an upper jawbone. Hypothesizing that eardrum evolution was related to these different jawbones, they performed a series of experiments that manipulated lower jaw development in mice and chickens.
First they examined eardrum development in mice that lacked the Ednra receptor, a condition known to inhibit lower jaw development. They found that these mice also lacked eardrums and ear canals, showing that their development was contingent on lower jaw formation.
Next, they used an Ednra-receptor antagonist to block proper development of the lower jaw in chickens. Rather than losing the eardrum, this manipulation created duplicate eardrums and ear canals, with the additional set forming from upper jaw components that had developed within the malformed lower jaw.
To understand how the eardrum evolved twice and why it is associated with different jaw components, the researchers looked at expression of Bapxl—a marker for the primary jaw joint—and its position relative to the first pharyngeal pouch. They found that in mouse embryos, Bapxl was expressed in cells slightly below the first pharyngeal pouch and that in chickens it was expressed considerably lower. This difference forces the eardrum to develop below the primary jaw joint in mammals, necessitating an association with the lower jaw, and above the joint in diapsids, necessitating an association with the upper jaw.
While scientists still do not know how or why the primary jaw junction shifted upwards in mammals, the study shows that the middle ear developed after this shift and must therefore have occurred independently after mammal and diapsid lineages diverged from their common ancestor. Emphasizing the importance of this evo-devo approach, Chief Scientist Shigeru Kuratani notes that, “convergent evolution can often result in structures that resemble each other so much that they appear to be homologous. But, developmental analyses can often reveal their different origins.”
For structures like the eardrum that do not fossilize, the evo-devo approach is even more important. Lead author Masaki Takechi speculates that, “this approach to studying middle ear evolution could help us understand other related evolutionary changes in mammals, including the ability to detect higher toned sounds and even our greater metabolic efficiency.”
Reference
- Taro Kitazawa, Masaki Takechi, Tatsuya Hirasawa et al., "Developmental genetic bases behind the independent origin of the tympanic membrane in mammals and diapsids", Nature Communications, doi: 10.1038/ncomms7853
Contact
Chief Scientist
Shigeru Kuratani
Evolutionary Morphology Laboratory
Chief Scientist Laboratories
Adam Phillips
RIKEN Global Relations and Research Coordination Office
Tel: +81-(0)48-462-1225 / Fax: +81-(0)48-463-3687
Email: pr@riken.jp
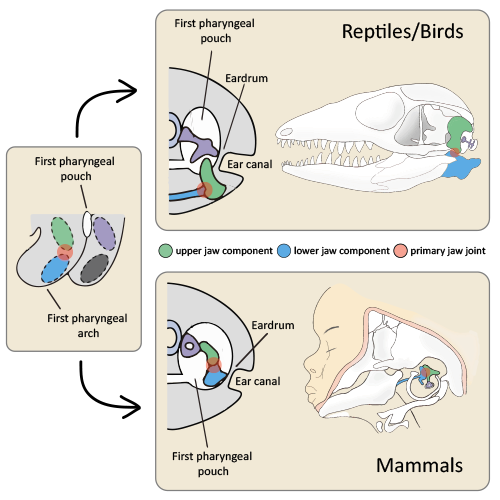
Morphology of the primary jaw joint in mammals and reptiles/birds
(Left) Generalized schematic showing the relative position of the first pharyngeal pouch and elements in the first pharyngeal arch. The upper (green) and lower (blue) elements in the first pharyngeal arch form the primary jaw joint (red circle). (Top) Reptiles/birds. The schematic on the left shows the eardrum forming between the first pharyngeal pouch and the ear canal. It is connected to one middle ear bone and the quadrate bone (green), which is part of the upper jaw. The schematic on the right shows these bones in a lizard with the primary jaw joint located between the upper and lower jaws. (Bottom) Mammals. The schematic on the left shows the eardrum forming between the first pharyngeal pouch and the ear canal. It is connected to the hammer bone (blue), which along with the anvil (green) make up 2 of the three middle ear bones. The schematic on the right shows the middle ear in a human embryo, with the primary jaw joint located within the middle ear, and the hammer (blue) connected to the lower jaw.
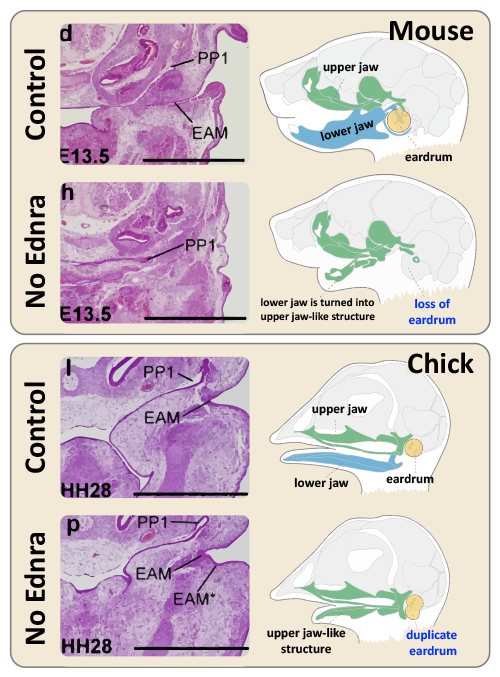
The effects of inhibiting Ednra in mice and chicks
(Top) The eardrum depends on lower jaw development in mice. The top panel shows the control condition. Histology shows a normal ear canal (external auditory meatus; EAM) and the first pharyngeal pouch (PP1). The diagram on the right shows a normal eardrum connected to the lower jaw. The bottom panel shows the effect of inhibiting Ednra, which disturbs the development of the lower jaw. The ear canal is missing in the histology, and the diagram shows how the lower jaw is transformed into an upper jaw-like structure. (Bottom) The eardrum is associated with upper jaw development in chicks. The upper panel shows a control case with a normal ear canal and an eardrum connected to the upper jaw. After inhibiting Ednra, histology shows the formation of a duplicate ear canal. The duplicate eardrum and lower jaw transformation are depicted in the schematic on the right.
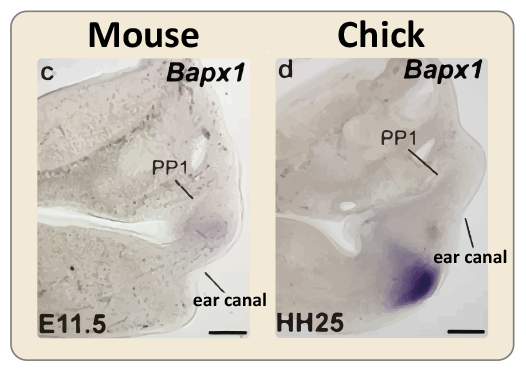
The primary joint develops more dorsally in mammals than in dyapsids
(Left) Bapx1 (purple) is a marker for the primary jaw joint. Early in mouse development, Bapx1 expression is very close to the first pharyngeal pouch (PP1), and the ear canal forms below it. (Right) In chicks, Bapx1 is expressed more ventrally, and the ear canal forms above it. Note that the relative position of PP1 and the ear canal are similar in mice and chicks.