2015年4月22日
理化学研究所
東京大学大学院医学系研究科
哺乳類と爬虫類-鳥類は、独自に鼓膜を獲得
-2億年以上前の進化の痕跡を発生学実験で明らかに-
要旨
理化学研究所(理研)倉谷形態進化研究室の倉谷滋主任研究員、武智正樹元研究員、東京大学大学院医学系研究科の栗原裕基教授、北沢太郎元大学院生らの共同研究グループ※は、マウスとニワトリの胚発生において同じ遺伝子の働きを抑える実験を行い、進化の中で哺乳類系統[1]と爬虫(はちゅう)類-鳥類系統[2]がそれぞれ独自の発生メカニズムにより鼓膜を獲得したことの発生学的証拠を発見しました。
陸上脊椎動物は、空気中の音を聴くために、鼓膜[3]と中耳骨[4]を顎(がく)関節の近くに進化させてきました。中耳骨は、哺乳類では3個、爬虫類と鳥類では1個あります。これらの骨は化石にも残ることから、その進化の歴史をたどることができ、哺乳類の祖先で顎とその支持装置を構成していた骨が次第に中耳の骨へと変化していった様子が明らかになっています。しかし、どのようなきっかけで、哺乳類系統が爬虫類-鳥類系統よりも多くの中耳骨を持つようになったのかは不明でした。また、鼓膜のような軟組織は化石には残らないため、鼓膜がいつ獲得されたのかは分かっていませんでした。
共同研究グループは、胚発生時に顎の前駆組織を下顎へ分化させる遺伝子の働きを抑える操作を行い、下顎の位置にも上顎の骨が発生するようなマウスとニワトリを作り出しました。マウスでは遺伝子ノックアウト技術[5]を使い、ニワトリでは発生中の胚に薬剤を投与して得たもので、骨格の形態に対してまったく同じ効果が出ることが確認できました。しかし、鼓膜に対しては、マウスとニワトリで正反対の影響が見られ、マウスでは鼓膜がなくなり、逆にニワトリでは鼓膜が上下に拡張することが分かりました。このことは、発生においてマウスの鼓膜は下顎の一部として、これに対しニワトリの鼓膜は上顎の一部として、つまり両動物が別々の発生メカニズムにより鼓膜を作っていることを示しています。
この発生メカニズムの明確な違いは、哺乳類系統と爬虫類-鳥類系統が分かれた後に、鼓膜が両系統で独立に獲得されたことを示唆します。また、鼓膜の位置の違いが、2系統の動物で異なる数の中耳骨を進化させたきっかけの1つであったと推測されます。
本研究は、英国のオンライン科学雑誌『Nature Communications』(4月22日付け)に掲載されます。
※共同研究グループ
理化学研究所 倉谷形態進化研究室
主任研究員 倉谷 滋 (くらたに しげる)
元研究員 武智 正樹 (たけち まさき)(現 東京医科歯科大学 大学院医歯学総合研究科 分子発生学分野 テニュアトラック助教)
東京大学 大学院医学系研究科 分子細胞生物学専攻 生化学・分子生物学講座
教授 栗原 裕基 (くりはら ひろき)
元大学院生 北沢 太郎 (きたざわ たろう)(現 日本学術振興会特別研究員 PD 疾患生命工学センター 動物資源学部門)
背景
陸上脊椎動物は、空気中の音の振動を鼓膜で受け取り、中耳骨を通して内耳の神経へ伝達します。中耳骨は、哺乳類では3個、爬虫類-鳥類では1個あります(図1)。化石種も含めた骨格の形態を系統図上で比較すると、これらの中耳骨は、祖先動物で顎の関節を構成していた骨から進化してきたことが分かります。ある骨がまったく別の機能を持つようになった進化過程を鮮やかに復元したこの一連の研究は「比較形態学の金字塔」と言えるものです。
哺乳類の中耳骨は、ツチ骨、キヌタ骨、アブミ骨からなり、祖先では顎の関節とその支持装置として機能していました。一方、爬虫類と鳥類における中耳骨(耳小柱)は哺乳類のアブミ骨と同じものですが、他の2つの骨はありません(図1)。なぜ、このように2つの進化パターンが起こったのか、何が進化の分かれ道になったのかは分かっていませんでした。
これについては2つのシナリオが考えられました。1つ目は、共通祖先で鼓膜が獲得された後に哺乳類系統と爬虫類-鳥類系統が分岐して異なる中耳の進化を遂げたというシナリオ、もう1つはそもそも両系統が分岐した後で独立に鼓膜が獲得されたというシナリオです。鼓膜を含めた軟組織そのものは化石に残らないため、化石種の研究からこれを明らかにするのには限界があり、別の証拠が必要とされていました。
共同研究グループは、鼓膜の発生メカニズムについて、特に周囲の骨格との結びつきを解明することを目的とし、マウスとニワトリをそれぞれ哺乳類系統と爬虫類-鳥類系統のモデル動物として下顎の特徴を上顎に変化させる実験を行い、鼓膜にどのような影響が生じるかを調べました。
研究手法と成果
脊椎動物の顎および鰓(えら)の骨格は、その前駆組織から上顎や下顎に分化していく過程でDlx遺伝子群[6]の制御を受けます。このうち、Dlx5/Dlx6遺伝子は前駆組織の腹側(下側)で発現し、その領域で下顎の骨要素が形成されます。対して、Dlx5/Dlx6遺伝子の発現がない背側(上側)の領域では、上顎の骨要素が形成されます。Dlx5/Dlx6遺伝子の働きを抑制すると、前駆組織の腹側領域も上顎として発生することが知られています。
共同研究グループは、マウスとニワトリに対して、このDlx5/Dlx6遺伝子を制御しているエンドセリン1シグナル伝達[7]に操作を加えて、下顎の特徴を上顎に変化させる実験を行いました。
マウスに対しては、遺伝子ノックアウト技術を施してエンドセリンA受容体を欠損した個体を作り出しました。一方、遺伝子操作が難しいニワトリに対しては、エンドセリンと受容体の結合を阻害するボセンタンという薬剤を胚発生時に投与し、エンドセリン1シグナル伝達を抑制しました。ボセンタン投与後のニワトリ胚を調べたところ、Dlx5/Dlx6遺伝子の発現が抑えられており、マウスとニワトリに施したこれらの実験操作はDlx5/Dlx6遺伝子に対して同様の効果を与えることが確認できました。この効果は骨格要素の形の変化としても表れ、実験操作を施したマウスとニワトリでは、本来は下顎の骨要素が占める位置に上顎の骨要素が形成されました(図2)。
ところが、興味深いことに、実験操作を施したマウスとニワトリでは、鼓膜に関して正反対の影響が見られました(図2)。マウスでは鼓膜がなくなり、ニワトリでは鼓膜のサイズが大きくなったのです。ニワトリの実験で見られた鼓膜の拡大について、その形態を正常個体と比較すると、背腹方向(上下方向)に重複したものであると確認できました。この結果は、マウスでは下顎ができる領域で鼓膜が発生し、ニワトリでは上顎ができる領域で鼓膜が発生することを示しています。
さらに共同研究グループは、マウスとニワトリにおける鼓膜の発生領域の違いに関して、より詳細な解析を進めました。下顎が分化するDlx5/Dlx6遺伝子発現領域の一部分では、Dlx5/Dlx6遺伝子によって制御されるGsc(グースコイド)遺伝子[8]が発現しています。胚の組織切片を作製し、Gsc遺伝子の発現を観察したところ、マウスとニワトリで同じ骨要素に発現が見られたものの、マウスではGsc遺伝子が発現している腹側(下側)の領域に鼓膜が形成され、ニワトリではGsc遺伝子が発現していない背側(上側)の領域で鼓膜が形成されることが分かりました。次に、上顎と下顎の関節が発生する位置について詳細に観察を行い、鼓膜の発生位置に対して、マウスではニワトリよりも関節が相対的に背側(上側)にできることを見いだしました。
これらの研究結果から、鼓膜の発生はマウスでは下顎に、ニワトリでは上顎に付随して生じることが分子発生学レベルで示されました。したがって鼓膜は共通祖先で獲得されたものではなく、哺乳類系統ではエンドセリン1シグナルに制御される下顎領域に鼓膜を獲得し、それとは別に、爬虫類-鳥類系統では上顎領域に鼓膜を獲得し、両系統でそれぞれ鼓膜と顎の位置関係を保持してきたという進化過程が強く支持されます(図3)。また、顎関節の位置のわずかな違いが進化の分かれ道となり、その後の両系統における中耳や顎の骨の進化パターンに影響したことも示唆しています(図3)。哺乳類系統で中耳の骨の1つへ進化した関節骨は下顎の骨に当たります(図1)。哺乳類系統では、最初に鼓膜が獲得された位置が下顎側であったために、下顎の骨とそれに関節する骨(方形骨)も鼓膜から内耳の神経に音を伝達する骨として進化したと考えられます(図1)。
今後の期待
今回の成果は、鼓膜という哺乳類系統と爬虫類-鳥類系統でそっくりな構造が、互いに独立に獲得されたことの痕跡が、発生メカニズムの違いとして残っていることを示唆したものです。特に哺乳類系統で、もともと下顎の骨だったものが中耳の骨へ組み込まれた進化は重要なステップであり、本研究成果をもとに、哺乳類が独自に獲得した聴覚や咀嚼(そしゃく)機能の進化についても、発生学的なアプローチからより詳細な研究が進んでいくと予想されます。
また、本研究のように、発生メカニズムに実験操作を施すことで、これまで認識されていなかった進化過程の痕跡が浮かび上がってくる例は、他にもありうると考えられます。私たちの体がなぜこのような形をしているのか、という問いに対して、今後、進化の歴史から掘り起こす本研究のようなアプローチはますます重要になっていくと期待できます。
原論文情報
- Taro Kitazawa, Masaki Takechi, Tatsuya Hirasawa et al., "Developmental genetic bases behind the independent origin of the tympanic membrane in mammals and diapsids", Nature Communications, doi: 10.1038/ncomms7853
発表者
理化学研究所
主任研究員研究室 倉谷形態進化研究室
主任研究員 倉谷 滋(くらたに しげる)
元研究員 武智 正樹(たけち まさき)(現 東京医科歯科大学 大学院医歯学総合研究科 分子発生学分野 テニュアトラック助教)
東京大学 大学院医学系研究科 分子細胞生物学専攻 生化学・分子生物学講座
教授 栗原 裕基(くりはら ひろき)
元大学院生 北沢 太郎(きたざわ たろう)(現 日本学術振興会特別研究員 PD 疾患生命工学センター 動物資源学部門)
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.哺乳類系統
哺乳類に至る系統は、約3億1300万年前に爬虫類-鳥類系統と分岐した。初期の仲間は現在の哺乳類とはかけ離れた姿をしており、約2億5000万年前から2億2500万年前にかけて顎の関節を含む哺乳類の特徴が進化してきた。私たちヒトを含む胎盤を持つ哺乳類の系統は、約1億2500万年前に出現した。 - 2.爬虫類-鳥類系統
現在生息している爬虫類と鳥類は、双弓類(そうきゅうるい)というグループに属している。この双弓類のうち、約1億5000万年前に恐竜から進化してきたのが鳥類である。なお、現生の爬虫類のうち、鳥類に最も近縁なのはワニである。 - 3.鼓膜
空気中の音の振動をとらえる薄い膜状の組織。鼓膜でとらえられた振動は中耳の骨へ伝達され、内耳で電気信号に変換され神経に伝わる。 - 4.中耳骨
哺乳類の中耳は、内耳側から鼓膜側に向かってアブミ骨、キヌタ骨、ツチ骨、が連なった構造を持つ。これらの骨は、それぞれ祖先動物の舌顎骨(ぜつがくこつ。上顎と他の頭骨をつなぐ骨)、方形骨(上顎の骨)、関節骨(下顎の骨)から進化した。爬虫類と鳥類における中耳骨は哺乳類のアブミ骨と同じものだが、他の2つの骨はない。 - 5.遺伝子ノックアウト技術
遺伝子組み換え操作によって、特定の遺伝子の働きを人為的に欠損させた動物を作製する技術。本研究では、まずエンドセリン1の受容体の1つであるエンドセリンA受容体遺伝子を欠損するように設計されたDNA配列を準備した。次にこの配列をゲノムに組み込んだマウス細胞を作製した。この細胞をマウス初期胚に移植し、出生した個体を用いて交配を重ねることにより、エンドセリンA受容体遺伝子を欠損するが、他の遺伝子機能は正常のままである遺伝子組み換えマウスを得た。 - 6.Dlx遺伝子群
脊椎動物の頭頸部の発生において重要な役割を果たす遺伝子群で、転写因子として顎の発生プログラムに必要な下流の遺伝子を制御する。脊椎動物の上顎の前駆組織には Dlx1/Dlx2遺伝子のみが発現するが、下顎の前駆組織には Dlx5/Dlx6遺伝子の発現も加わるという入れ子状の発現パターンを示し、これは Dlxコードとも呼ばれる。 Dlx5/Dlx6遺伝子を不活性化すると下顎と上顎の Dlxコードが同じになり、下顎の形態が上顎のそれに変化する。 - 7.エンドセリン1シグナル伝達
エンドセリン1は21個のアミノ酸からなる小さなペプチドであり、産生細胞から分泌され、その受容体を発現する細胞に働きかけることでシグナルを伝達する。当初は血管収縮因子として発見されたが、発生過程においては下顎の前駆組織にエンドセリンA受容体を通して働きかけて Dlx5/Dlx6遺伝子の発現を誘導することが知られている。実際、エンドセリン1のシグナル伝達を胚発生の途中で阻害すると Dlx5/Dlx6遺伝子の発現が抑制され、 Dlx5/Dlx6遺伝子を不活性化させた時と同様に下顎が上顎に変化する。 - 8.Gsc(グースコイド)遺伝子
Dlx5/Dlx6遺伝子の下流で働く転写因子であり、下顎の一部に発現し、その発生を制御する。
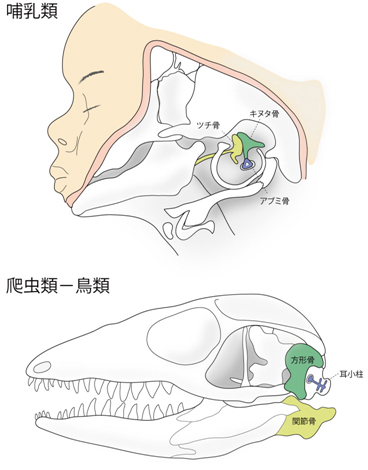
図1 哺乳類(ヒト胎児)と爬虫類-鳥類(トカゲ)の中耳の構造
哺乳類はツチ骨(黄)、キヌタ骨(緑)、アブミ骨(青)の3つの中耳骨を有するが、爬虫類-鳥類にはアブミ骨に相当する骨要素(耳小柱)しか存在しない。哺乳類の他の2つの骨要素は、祖先動物において顎の関節を構成していた関節骨と方形骨から進化した。一方、爬虫類-鳥類では、顎の関節は祖先動物から大きく変化しておらず、関節骨(黄)と方形骨(緑)の間にある。
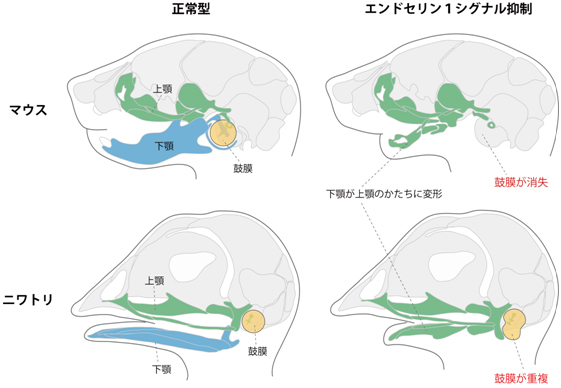
図2 正常型(左)と実験操作を施した(右)マウスとニワトリ
実験操作により胚発生時のエンドセリン1シグナル伝達を抑制したマウスとニワトリでは、ともに、本来は下顎の骨要素(青色)が占める位置に上顎の骨要素(緑色)が形成される。一方、この実験操作は鼓膜に対してはマウスとニワトリでは正反対の影響を及ぼし、マウスでは鼓膜がなくなり、ニワトリでは鼓膜が上下に重複した。
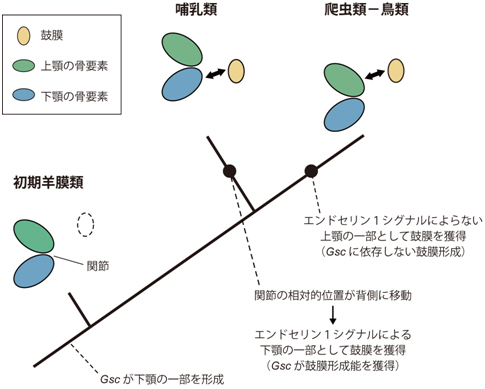
図3 本研究に基づいた鼓膜と中耳の進化過程
哺乳類と爬虫類-鳥類の共通祖先である初期羊膜類には鼓膜は存在せず、エンドセリン1シグナル伝達の一部であるGsc遺伝子は下顎の形成に関わっていた。哺乳類では鼓膜に対する顎関節の相対的位置が背側(上側)に移動した結果、Gsc遺伝子が鼓膜形成能を獲得し、下顎の一部として鼓膜を獲得したが、爬虫類-鳥類系統では上顎の一部として鼓膜が獲得された。