理化学研究所(理研)生命機能科学研究センター細胞外環境研究チームのチュンチュン・チェン研究員(研究当時)、筒井仰研究員(研究当時)、藤原裕展チームリーダー、新潟医療福祉大学の田口徹教授らの国際共同研究グループ※は、マウス毛包[1]の「表皮幹細胞[2]」が触覚受容器の正常な機能に必須であることを突き止め、皮膚で触覚が生まれる仕組みの一端を解明しました。
本研究成果は、表皮幹細胞は皮膚の再生だけでなく、触覚受容に欠かせない感覚神経終末[3]の形と機能を制御するという触覚機構の新しいメカニズムの発見であり、皮膚の触覚機構の再生や老年期の触覚機能の低下を防ぐ方法の開発などに貢献すると期待されます。
触覚は、五感のうち最も原始的な感覚で、生物の生存に不可欠です。皮膚には感覚神経の終末などからなる「触覚受容器」があり、それがさまざまな物理刺激を化学シグナルに変換した情報を、感覚神経を通して中枢神経系に伝えることで触覚を生み出しています。しかし、皮膚の触覚受容器が作られ、機能する仕組みはよく分かっていませんでした。
今回、国際共同研究グループは、毛包と毛の再生を担う表皮幹細胞が、感覚神経終末の周囲に特殊な細胞外マトリックス[4]を形成することで、感覚神経と一体となって触覚受容器の複雑な構造と機能を生み出していることを明らかにしました。また、表皮幹細胞が、ダイナミックに再生を繰り返す毛包の中に安定かつ恒常的な組織構造を作り出すことで、毛包と感覚神経が安定に接続されることも明らかにしました。
本研究は、英国の科学雑誌『eLife』(10月25日付け)に掲載されました。
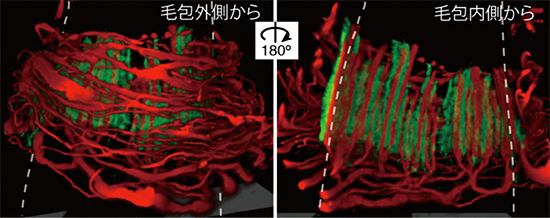
図 毛包表皮幹細胞の細胞外マトリックス(緑)が、触覚受容器の感覚神経(赤)のパターンを制御する
※国際共同研究グループ
理化学研究所 生命機能科学研究センター
細胞外環境研究チーム
研究員(研究当時) チュンチュン・チェン(Chun-Chun Cheng)
研究員(研究当時) 筒井 仰(つつい こう)
テクニカルスタッフII 伴(三千) 典子(ばん(さんぜん) のりこ)
テクニカルスタッフII 中川 麻子(なかがわ あさこ)
チームリーダー 藤原 裕展(ふじわら ひろのぶ)
超微形態研究チーム
テクニカルスタッフI(研究当時) 垣口 貴沙(かきぐちきさ)
チームリーダー 米村 重信(よねむらしげのぶ)
分子配列比較解析ユニット
テクニカルスタッフI(研究当時) ショーン・D・キーリー(Sean D. Keeley)
テクニカルスタッフII 種子島 千春(たねがしま ちはる)
生体モデル開発ユニット
ユニットリーダー 清成 寛(きよなり ひろし)
生体ゲノム工学研究チーム
チームリーダー 古田 泰秀(ふるた やすひで)
新潟医療福祉大学 理学療法学科
教授 田口 徹(たぐち とおる)
(研究当時:名古屋大学環境医学研究所助教)
重井医学研究所分 子細胞生物部門
部長 友野 靖子(ともの やすこ)
キングス・カレッジ・ロンドン
教授 フィオナ・M・ワット(Fiona M. Watt)
※研究支援
本研究は、日本学術振興会(JSPS)科学研究費補助金新学術領域(研究領域提案型)「領域特殊化された基底膜が仲介する知覚神経ー毛包幹細胞相互依存(研究代表者:藤原裕展)」、上原記念生命科学財団「基底膜が異種組織を繋ぐ機構(研究代表者:藤原裕展)」、武田科学振興財団「基底膜による毛包幹細胞ー感覚神経相互依存性の制御機構(研究代表者:藤原裕展)」、コスメトロジー研究振興財団「皮膚感覚神経ー毛包幹細胞相互依存における基底膜領域特殊化の役割(研究代表者:藤原裕展)」などの支援を受けて行われました。
背景
触覚は五感のうち最も原始的な感覚であり、外界を感知する基本機能として生物の生存に不可欠です。例えば、ヒトの皮膚は、体表のあらゆる部位に幾万もの「触覚受容器」を配置している人体最大の感覚器官です。触覚受容器は、体表で生じるさまざまな物理刺激を化学シグナルに変換し、その情報を、感覚神経を介して中枢神経系に伝えることで触覚を生み出しています。触覚には、対象に接触しなければ感覚を得られないという欠点がありますが、多くの動物は、表皮細胞から毛や羽などを作ることで、外界から体を保護しつつ、知覚可能な空間領域を拡張しています。毛は皮膚表面から突出したセンサープローブとして機能し、そこから生じる物理刺激を、毛を作る器官である毛包内の触覚受容器に伝達します(図1A)。触覚受容器を備えた毛包はさまざまな動物で見られることから、毛包は進化的に保存された触覚器官といえます。
毛包は再生する器官であり、胎児期に形成された後も、一生を通じて退行・休止(脱毛)・成長を繰り返す再生周期(毛周期[1])を持ちます。そのため毛包には、再生を担う表皮幹細胞が、毛包の少し膨らんだバルジと呼ばれる領域に常在します。最近の研究により、表皮幹細胞の集団は均一ではなく多様性があること、そして異なる性質の幹細胞が表皮内で区画化され、まとまったコンパートメントを形成していることが分かってきました。しかし、なぜ毛を作るための表皮幹細胞が多様性を獲得し、区画化されるのかは十分理解されていませんでした。さらに、毛包内の触覚受容器は、感覚神経終末、シュワン細胞、表皮細胞、そして細胞外マトリックスなどからなる複雑な構造を持ちますが、これがどのように形成され、毛周期を通じて機能が維持されるかについての詳しい仕組みも分かっていませんでした。
2011年に国際共同研究グループはマウスを用いた研究から、バルジ中央部の表皮幹細胞が特定の細胞外マトリックスタンパク質を分泌することで、近傍に位置する立毛筋(鳥肌を立てるために必要な筋肉)の発生と、その毛包への接続を制御することを見いだしていました注1)。一方、触覚受容器の神経終末は、立毛筋より少し上のバルジ上部に接続し、バルジ上部の幹細胞と近接しています。このことから、毛包の表皮幹細胞は位置によって異なる役割を担っており、バルジ上部の幹細胞は毛包の再生だけではなく、触覚受容器の形成と機能にも寄与するという仮説を立て、その検証を試みました。
注1)Fujiwara et al., 2011. The basement membrane of hair follicle stem cells is a muscle cell niche. Cell. 144, 577-589
研究手法と成果
国際共同研究グループはまず、マウス毛包の表皮幹細胞を、毛包内での位置と発現する遺伝子の違いをもとに五つの亜集団に分類し、各幹細胞集団をセルソーター[5]で純度高く分離できる方法を確立しました。次に、これらの細胞集団の詳細な遺伝子発現プロファイルを、RNAシーケンス法[6]により解析しました。すると、触覚受容器の神経終末と接しているバルジ上部の表皮幹細胞では、神経誘引因子などの神経形成に関わる遺伝子が強く発現していることが分かりました。さらに、他の幹細胞集団とは異なる細胞外マトリックス遺伝子を多数発現していることも分かりました。
そこで、これらの遺伝子から作られるタンパク質の組織内での局在を調べたところ、EGFL6[7]と呼ばれる細胞外マトリックスタンパク質が、触覚受容器の神経終末の周囲に沈着していることが分かりました(図1B)。電子顕微鏡による超微細構造の解析から、触覚受容器の周囲には、EGFL6を含む襟状の細胞外マトリックスが存在することを見いだし、国際共同研究グループはこの新しい細胞外マトリックス構造を「カラー(襟)マトリックス」(collar matrix)と名付けました。
カラーマトリックスの役割を調べるため、Egfl6遺伝子を欠損したマウスの神経終末を蛍光顕微鏡で観察し、その構造に生じた異常を3次元的に定量解析しました。EGFL6タンパク質を欠失した毛包では、毛包の長軸と平行に配置されたストライプ状の神経終末のパターンが乱れ、隣の神経終末と接触する頻度が増えることが分かりました(図2A)。さらに、この毛包の触覚機能を調べたところ、触刺激に対する毛包由来感覚神経の応答(活動電位数)が野生型に比べて大きく減少し、特に弱い刺激に対する反応が低下していました(図2B)。
以上の結果から、表皮幹細胞から分泌されるEGFL6タンパク質が、触覚受容器の構造と触覚機能に重要な役割を果たしていることが明らかとなりました。哺乳類の触覚受容器で機能する細胞外マトリックスの発見は、本研究が初めてです。
続いて、毛包組織が消長する毛周期の過程で、毛包と神経終末との安定的な接続が維持される仕組みを調べるため、毛周期でのバルジ組織の構造変化に着目しました。マウスの体毛では、胎仔期に形成されたバルジの表皮幹細胞は、出生後の毛周期ではほとんど利用されることなくそのまま静止状態を保ちます。代わりに、一部の表皮幹細胞が分裂して毛包の頭尾軸の頭側に新たなバルジと毛包を形成し、そこから新しい毛を産生します。新しいバルジと古いバルジが頭尾方向に直列につながった構造は、ダブル・バルジと呼ばれます(図3)。
国際共同研究グループは、毛周期でのダブル・バルジ構造と神経終末、カラーマトリックスの関係を詳しく解析した結果、以下のことを見いだしました。
- 1.新しいバルジが毛包頭部側にできるため、頭部側の表皮構造はダイナミックに変化するが、尾部側の表皮(古いバルジ)は、組織構造をほとんど変えない(図4左)
- 2.新たなバルジができバルジの周囲長が2倍になると、バルジ周囲をリング状に取り囲んでいた神経終末と細胞外マトリックス(EGFL6タンパク質)の構造が、毛包尾部側に偏ったコの字型に変化する(図4左)。
- 3.古いバルジの幹細胞を人為的に除去すると、バルジ尾部側の表皮構造が変化するとともに、神経終末がバルジ頭部側(本来は神経終末が存在しない)に移動する(図4右)。
これらの結果から、古いバルジにある静止状態の表皮幹細胞が、毛周期のステージにかかわらず毛包尾部側での毛包と神経終末との安定的な接続点を維持する働きを持つことが示されました。尾部側に偏った神経終末の分布は、毛の頭尾方向の揺れ方向を検知するために必要であることが最近報告されています注2)。よって本研究で示された、頭尾方向に極性のあるダブル・バルジ構造が神経終末をリング状からコの字型に変化させる働きは、毛周期を通じて毛の揺れ方向を安定的に感知するための重要な仕組みであると言えます。
注2)Rutlin et al., 2014. The cellular and molecular basis of direction selectivity of Ad-LTMRs. Cell 159, 1640-1651.
今後の期待
これまでの触覚研究は、感覚神経自体に主眼をおいたものがほとんどでしたが、本研究により、その周囲に存在する非神経系の表皮幹細胞や細胞外マトリックスが触覚機能に極めて重要であることが示されました。細胞外マトリックスが、物理刺激の化学シグナルへの変換にどのように関与しているのかは今後の重要な課題です。加齢に伴う触覚機能の低下についても、神経自体ではなく、表皮幹細胞やマトリックスの加齢変化が関与している可能性があります。今後、非神経系の要素をターゲットとした触覚機能の発生・再生研究やその医療応用が進むと考えられます。
また今回の研究は、表皮幹細胞の多様性と区画化の意義についても新たな視点を与えています。表皮幹細胞は表皮組織の維持と再生を担うだけではなく、さまざまな性質の幹細胞を皮膚の適切な位置に配置し、周囲の細胞と相互作用を成立させることで、器官の恒常性と高次機能の発現を両立させていることが分かりました。異なる組織や臓器間の協調関係(チームワーク)に目を向けることで、細胞社会の秩序を生み出す仕組みが今後も明らかになると期待されます。
原論文情報
- Chun-Chun Cheng, Ko Tsutsui, Toru Taguchi, Noriko Sanzen, Asako Nakagawa, Kisa Kakiguchi, Shigenobu Yonemura, Chiharu Tanegashima, Sean D. Keeley, Hiroshi Kiyonari, Yasuhide Furuta, Yasuko Tomono, Fiona M. Watt and Hironobu Fujiwara, "Hair follicle epidermal stem cells define a niche for tactile sensation", eLife, 10.7554/eLife.38883
発表者
理化学研究所
生命機能科学研究センター 細胞外環境研究チーム
チームリーダー 藤原 裕展(ふじわら ひろのぶ)
研究員(研究当時) チュンチュン・チェン(Chun-Chun Cheng)
(現 UT Southwestern Medical Center 研究員)
研究員(研究当時) 筒井 仰(つつい こう)
(現 テクニカルスタッフI)
新潟医療福祉大学 理学療法学科
教授 田口 徹(たぐち とおる)
(研究当時:名古屋大学 環境医学研究所 助教)
 藤原 裕展
藤原 裕展
 チュンチュン・チェン
チュンチュン・チェン
 筒井 仰
筒井 仰
 田口 徹
田口 徹
お問い合わせ先
理化学研究所 生命機能科学研究センター センター長室 報道担当
山岸 敦(やまぎし あつし)
Tel: 078-304-7138 / Fax: 078-304-7112
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
お問い合わせフォーム
新潟医療福祉大学 入試広報部 広報課
中原 英伸(なかはら ひでのぶ)
Tel: 025-257-4459
E-mail:nakahara[at]nuhw.ac.jp
産業利用に関するお問い合わせ
補足説明
- 1.毛包、毛周期
毛を取り囲む皮膚の付属器官を毛包と呼び、毛髪や体毛を産生する。胎児期に形成された毛包は、成長期、退行期、休止期の順に周期的な再生を繰り返す。これを毛周期と呼ぶ。ヒトの頭髪の場合、数年間の成長期の後、数週間をかけて退行期に移行し、数カ月の休止期を経て脱落する。 - 2.表皮幹細胞
皮膚の基底層に存在し、表皮のもととなる幹細胞。毛包を囲む基底層は表皮の基底層と連続しており、毛包の幹細胞は表皮幹細胞の一種である。 - 3.神経終末
神経の軸索の末端。他の神経細胞や筋肉細胞などとシナプスを形成する。 - 4.細胞外マトリックス
細胞と細胞の間を満たし、生体組織を包み込む高分子の構造体。多糖高分子やタンパク質などを主成分とする。皮膚の基底層を裏打ちする基底膜は、細胞外マトリックスの一種。 - 5.セルソーター
細胞分離装置の一つ。細胞集団の中から任意の特徴(大きさ、形態、細胞内成分など)を持つ細胞を自動的に分離する装置。 - 6.RNAシーケンス法
組織や細胞で発現している全RNA(トランスクリプトーム)を解析する手法の1つ。mRNAやncRNAの断片的な配列情報(約50-125塩基)を網羅的に取得し、ゲノム配列と対応させることで、遺伝子発現量の定量や新たな転写配列の発見を行う。 - 7.EGFL6
基底膜分子であるネフロネクチンファミリーに属する細胞外マトリックスタンパク質。
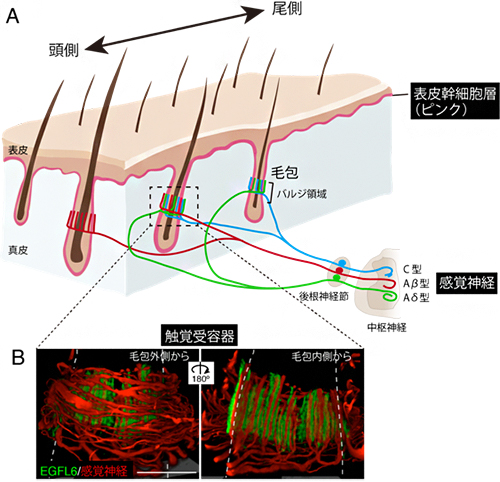
図1 皮膚毛包の触覚受容器
- (A) 皮膚の毛包のバルジと呼ばれる領域に触覚受容器が形成される。皮膚の表面で発生した物理的な刺激は、触覚受容器で化学シグナルに変換され、3種類の感覚神経(C型、Aβ型、Aδ型)を介して中枢神経系まで伝達される。
- (B) 触覚受容器の神経終末(赤)とEGFL6タンパク質(緑)の蛍光免疫組織像。破線は毛包の輪郭を表す。3次元観察により、毛包の外側(左)と内側(右)から見た状態を示した。EGFL6は表皮幹細胞から分泌される細胞外マトリックスタンパク質で、触覚受容器に特異的に存在する。スケールバーは20マイクロメートル(μm、1μmは1,000分の1mm)。
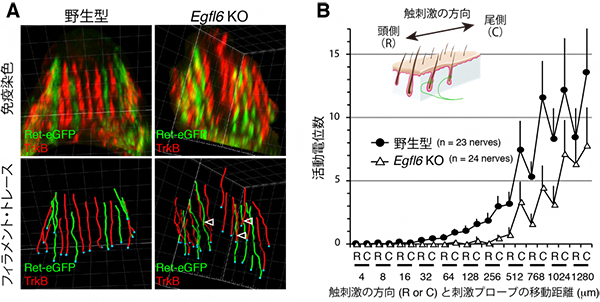
図2 Egfl6を欠損したマウスでは、触覚受容器が異常を呈する
- (A) 触覚受容器の感覚神経の免疫蛍光染色像(上)と、神経繊維の線描(下)。緑色はC型の、赤色はAδ型の神経繊維を示す。Egfl6欠損マウス(Egfl6 KO)では、神経終末の縦方向のストライプパターンが乱れ、隣の神経終末と重なる頻度が増える(矢頭)。白線格子の1マスの幅は5μm。
- (B) 皮膚のex vivo(生体外の)単一神経電気生理実験により、触覚受容器の応答特性を調べた。グラフは、刺激を与えるプローブ(針)の移動の方向(Rは頭側、Cは尾側)・距離(横軸)と、その時の活動電位の数(縦軸)を示したもの。Egfl6欠損マウス(Egfl6 KO)では、触刺激に対する感覚神経の応答(活動電位数)が有意に低下する。特に、グラフの真ん中から左側で見られるように、弱い触刺激で著しい。
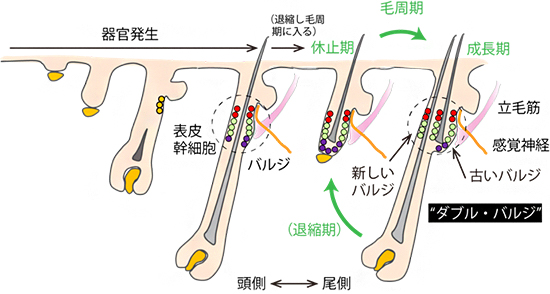
図3 毛周期とバルジの複製
毛包は、器官発生が完了するとアポトーシス(プログラムされた細胞死)により退縮し(休止期)、毛の伸長を停止する。その後、成長期、退縮期(図では省略)、休止期からなる毛周期を繰り返し、毛包組織と毛を再生する。マウスの毛周期では、毛包の表皮幹細胞は皮膚の頭尾軸の頭側に新たなバルジと毛包を形成し、尾側に残る古いバルジと直列に並んだダブル・バルジ構造をとる。毛周期を通じて触覚受容器の神経終末は、立毛筋より少し上のバルジ上部に接続し、バルジ上部の幹細胞(赤丸)と近接する。
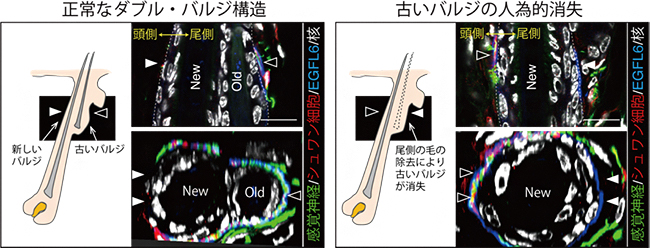
図4 表皮幹細胞による感覚神経パターンの制御
- (左) 正常なダブル・バルジでは、古いバルジ(old)構造は、毛周期を通して安定的に維持される。感覚神経(緑)とシュワン細胞(赤)、細胞外マトリックス(青)は尾側(old側)に偏って分布し、コの字状にダブル・バルジを取り囲む。
- (右) 尾側の毛を除去すると、古いバルジの表皮幹細胞にだけ特異的にアポトーシスが誘導され、古いバルジ構造が消失する。この時、感覚神経とシュワン細胞、細胞外マトリックスが頭側に移動する。黒矢頭は感覚神経の存在する領域、白矢頭は感覚神経の存在しない領域をそれぞれ示す。上段の顕微鏡像は毛包の垂直断面、下段は横断面。スケールバーは10μm。