理化学研究所(理研)環境資源科学研究センター適応制御研究ユニットの瀬尾光範ユニットリーダー、奈良先端科学技術大学院大学先端科学技術研究科バイオサイエンス領域の清水崇史助教(理研環境資源科学研究センター適応制御研究ユニット客員研究員)らの共同研究チームは、気孔[1]の閉鎖を誘導する植物ホルモン[2]であるアブシシン酸(ABA)[3]の輸送が、植物体内で制御される新たな仕組みを明らかにしました。
本研究成果は、気孔の開度を人為的に調節し、植物の乾燥耐性や生産性を向上させるための技術開発に貢献すると期待できます。
ABAは主に葉の維管束組織[4]の細胞で合成され、孔辺細胞[5]まで運ばれます。これまでに、ABAの細胞内への取り込み、もしくはABAの細胞外への排出を担うタンパク質(輸送体[6])が複数同定されてきましたが、植物体内でのABAの動きがどのように調節されているかは完全には理解されていませんでした。
今回、共同研究チームは、シロイヌナズナ[7]においてABA輸送体として機能することが報告されていたNPF4.6[8]タンパク質が、葉の孔辺細胞でABAの取り込みを行うことにより、気孔の閉鎖を促進することが分かりました。さらに、NPF5.1[8]と呼ばれる別のタンパク質がABA輸送体として機能することを明らかにしました。NPF5.1は葉の維管束組織や葉肉細胞[9]などで細胞内へのABAの取り込みを行うことで、維管束組織から孔辺細胞へと輸送されるABAの量を制限し、気孔の閉鎖を抑制すると考えられます。
本研究は、科学雑誌『Genes』オンライン版(6月8日付)に掲載されました。
背景
地上を自由に動き回ることができない植物には、環境の変化を耐え抜くためのさまざまな仕組みが備わっています。例えば、乾燥による土壌の水分不足に対応するために、葉の表面に存在する気孔を閉じることで、植物体からの水分の損失を防ぎます。一方で、光合成の効率は気孔からの二酸化炭素の取り込み量に依存するため、気孔が閉じることは自身の成長には不都合です。そのため、植物は多様な環境条件下で気孔の開き具合(開度)を適切に調節しています。
植物ホルモンの一つであるアブシシン酸(ABA)には、気孔を閉じる働きがあります。植物は、乾燥を感じるとABAを大量に合成し気孔を閉じます。そして乾燥が終わり再び水分が十分になると、過剰なABAが分解(不活性化)され気孔が開きます。
ABAは主に葉の維管束組織の細胞で合成され、孔辺細胞まで運ばれます。これまでに、ABAの細胞内への取り込み、もしくはABAの細胞外への排出を担うタンパク質(輸送体)が複数同定されてきましたが、植物体内でのABAの動きがどのように調節されているのかは完全には理解されていませんでした。
研究手法と成果
瀬尾光範ユニットリーダーらのこれまでの研究により、シロイヌナズナのNPF4.6タンパク質(NRT1.2もしくはAIT1とも呼ばれる)がABAを細胞内に取り込む輸送体として機能することが明らかになっていましたが注1)、葉の気孔開度に対するNPF4.6の働きは分かっていませんでした。
そこで共同研究チームは、気孔開度の間接的な指標となるシロイヌナズナの葉の表面温度[10]を調べました。すると、NPF4.6の機能を失ったnpf4.6変異体の葉の表面温度は野生型と同程度ですが、npf4.6変異体と内生ABA量が低下したaao3変異体を掛け合わせたaao3 npf4.6二重変異体では、aao3変異体に比べて葉の表面温度が低いことが分かりました(図1B)。これは、aao3 npf4.6二重変異体の気孔の方がaao3変異体よりも大きく開いていることを示しています。つまり、正常なNPF4.6には気孔の閉鎖を促進する働きがあると言えます。
また、NPF4.6タンパク質は葉の維管束組織および孔辺細胞に存在しますが(図2)、aao3 npf4.6二重変異体の孔辺細胞だけで正常なNPF4.6を発現させる(aaso3 npf4.6/pMYB60:NPF4.6)と、葉の表面温度がaao3 npf4.6二重変異体よりも上昇し、気孔の閉鎖が促進されることが分かりました(図1B)。このことから、NPF4.6は孔辺細胞内へのABAの取り込みを促進する因子であると考えられます。
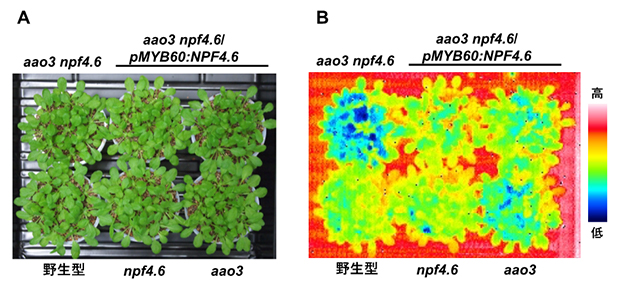
図1 野生型シロイヌナズナおよび各種変異体における葉の表面温度
- A)発芽後約1カ月の各種植物体。
- B)赤外線カメラ(サーモグラフィー)で観察した葉の表面温度。NPF4.6の機能を失ったnpf4.6変異体(下段中央)の葉の表面温度は野生型(下段左)と同程度だったが、二重変異体aao3 npf4.6(上段左)では、葉の表面温度がaao3変異体(下段右)に比べて低かった。また、上段中央・右のように、aao3 npf4.6二重変異体の孔辺細胞だけで正常なNPF4.6を発現させる(aaso3 npf4.6/pMYB60:NPF4.6)と、葉の表面温度が上昇した。
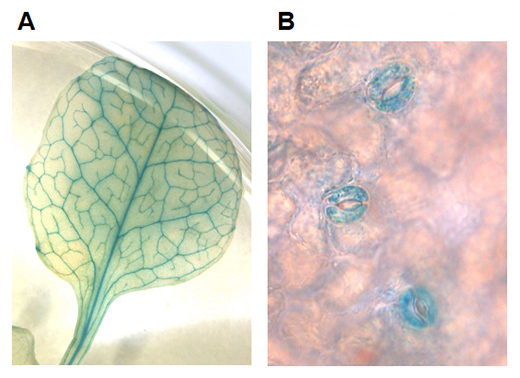
図2 シロイヌナズナにおけるNPF4.6タンパク質の局在部位
NPF4.6遺伝子プロモーター支配下でNPF4.6とレポーター(GUS)の融合タンパク質を発現させ、GUS染色によりNPF4.6タンパク質の局在部位(青く染まった部位)を可視化した。
- A)ロゼット葉の維管束組織でGUS活性が見られる。
- B)ロゼット葉表面の拡大写真。3組の孔辺細胞でGUS活性が見られる。
さらに、ABA取り込み活性を持つNPF5.1タンパク質の機能を失ったnpf5.1変異体では、野生型に比べて葉の表面温度が高いことが明らかになりました(図3B)。これは、npf5.1の気孔の開度が野生型に比べて小さいことを示しています。つまり、正常なNPF5.1には気孔の閉鎖を抑制する働きがあることを意味します。
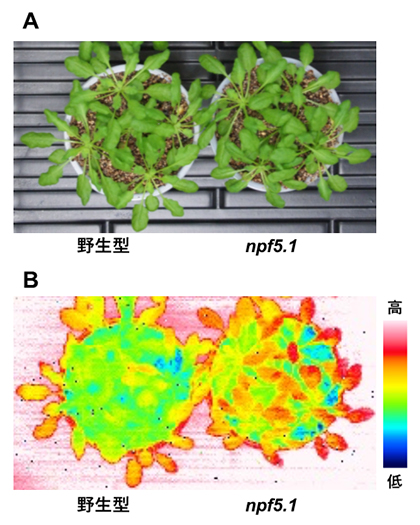
図3 野生型シロイヌナズナおよびnpf5.1変異体における葉の表面温度
- A)発芽後約1カ月の各種植物体。
- B)赤外線カメラ(サーモグラフィー)で観察した葉の表面温度。npf5.1変異体では、野生型に比べて葉の表面温度が高かった。
NPF5.1遺伝子の発現が葉の維管束組織や葉肉細胞などで見られることから(図4)、NPF5.1は生合成部位である維管束組織から孔辺細胞へ輸送されるABAの量を制限し、気孔の閉鎖を抑制すると考えられます(図5)。
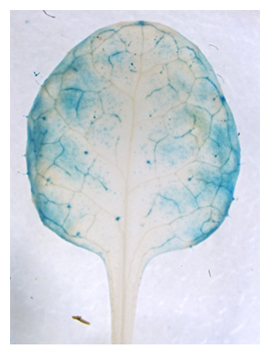
図4 NPF5.1遺伝子の発現部位
NPF5.1遺伝子プロモーター支配下でレポーター(GUS)を発現させ、GUS染色によりNPF5.1の発現部位(青く染まった部位)を可視化した。
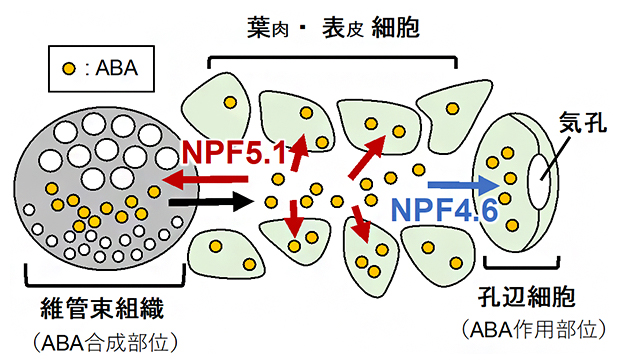
図5 NPF4.6およびNPF5.1によるABA輸送の制御モデル
NPF4.6は孔辺細胞でのABAの取り込みを介して気孔の閉鎖を促進する(青矢印)。NPF5.1は、維管束組織や葉肉細胞でのABAの取り込みを促進することで気孔の閉鎖を抑制する(赤矢印)。黒矢印は、他のABA輸送体の働きによるABA生合成部位(維管束組織)からのABAの排出を表す。
- 注1)2012年5月29日プレスリリース「栄養素を運ぶタンパク質「NRT1.2」が植物ホルモンも運ぶことを発見」
今後の期待
今回の研究で、細胞内にABAを取り込むという点においては同じ機能を持つ二つのABA輸送体が、その発現部位の違いによって気孔の閉鎖を促進することも抑制することも可能であることが示されました。この知見をもとに、ABA輸送体の発現部位を人為的にコントロールすることで、乾燥耐性や水利用効率の高い作物の作出が可能になるものと期待できます。
今回の研究成果は、国際連合が2016年に定めた17項目の「持続可能な開発目標(SGDs)[11]」のうち、「2.飢餓をゼロに」と「15.陸の豊かさを守ろう」に大きく貢献します。
補足説明
- 1.気孔
維管束植物の主に葉や茎の表面に存在する小さな孔。根から吸い上げた水分の蒸散や、光合成における二酸化炭素の吸収、酸素の排出に使われる。 - 2.植物ホルモン
植物体内で作られ、低濃度で作用し、植物種を超えて普遍的に存在する生理活性物質の総称。低分子性のものとしてはアブシシン酸のほかに、オーキシン、サイトカイニン、ジベレリン、エチレン、ブラシノステロイド、ジャスモン酸、サリチル酸、ストリゴラクトンなどが知られている。 - 3.アブシシン酸(ABA)
植物ホルモンの一つ。気孔の閉鎖を促進するほかに、ストレス応答性遺伝子の発現制御、種子休眠性の誘導といった生理作用を持つ。 - 4.維管束組織
道管や師管といった、水分や栄養分の通り道となる管状の構造を含む組織。 - 5.孔辺細胞
気孔を形成する二つで1組の細胞。 - 6.輸送体
細胞や細胞小器官(オルガネラ)を仕切る生体膜を介した物質の輸送をつかさどる、膜局在性のタンパク質。トランスポーター(transporter)とも呼ばれる。 - 7.シロイヌナズナ
アブラナ科の一年草。モデル実験植物としてさまざまな研究の材料として使われる。 - 8.NPF4.6、NPF5.1
どちらも、硝酸輸送体(NRT1)、ペプチド輸送体(PTR)を含むNRT1/PTR FAMILY(NPF)タンパク質の仲間。シロイヌナズナには53種類のNPFタンパク質が存在する。 - 9.葉肉細胞
葉の最外層に位置する細胞(表皮細胞)の内側に存在し、維管束組織に含まれない細胞。 - 10.葉の表面温度
気孔の開き具合(開度)を間接的に示す指標となる。すなわち、気孔が開くことで蒸散量が増加し、気化熱によって表面の温度が低下する。逆に、気孔が閉じることで蒸散量が低下し、表面温度が上昇する。 - 11.持続可能な開発目標(SGDs)
2015年9月の国連サミットで採択された「持続可能な開発のための2030アジェンダ」にて記載された2016年から2030年までの国際目標。持続可能な世界を実現するための17のゴールから構成され、地球上の誰一人として取り残さないことを誓っている。SDGsは発展途上国のみならず、先進国自身が取り組むユニバーサル(普遍的)なものであり、日本も積極的に取り組んでいる(外務省のホームページから一部改変して転載)。SDGs はsustainable development goalsの略。
共同研究チーム
理化学研究所 環境資源科学研究センター 適応制御研究ユニット
テクニカルスタッフⅡ 菅野 裕理(かんの ゆり)
特別研究員 鈴木 洋弥(すずき ひろみ)
基礎科学特別研究員 渡邊 俊介(わたなべ しゅんすけ)
ユニットリーダー 瀬尾 光範(せお みつのり)
奈良先端科学技術大学院大学 先端科学技術研究科先端科学技術研究科
バイオサイエンス領域
助教 清水 崇史(しみず たかふみ)
(理化学研究所 環境資源科学研究センター 適応制御研究ユニット 客員研究員)
原論文情報
- Takafumi Shimizu, Yuri Kanno, Hiromi Suzuki, Shunsuke Watanabe and Mitsunori Seo, "Arabidopsis NPF4.6 and NPF5.1 Control Leaf Stomatal Aperture by Regulating Abscisic Acid Transport", Genes, 10.3390/genes12060885
発表者
理化学研究所
環境資源科学研究センター 適応制御研究ユニット
ユニットリーダー 瀬尾 光範(せお みつのり)
奈良先端科学技術大学院大学 先端科学技術研究科先端科学技術研究科
バイオサイエンス領域
助教 清水 崇史(しみず たかふみ)
(理化学研究所 環境資源科学研究センター 適応制御研究ユニット 客員研究員)
 瀬尾 光範
瀬尾 光範
 清水 崇史
清水 崇史
報道担当
理化学研究所 広報室 報道担当
お問い合わせフォーム
奈良先端科学技術大学院大学 企画総務課 渉外企画係
Tel: 0743-72-5063 / Fax: 0743-72-5011
Email: s-kikaku [at] ad.naist.jp
※[at]は@に置き換えてください。