2009年7月10日
独立行政法人 理化学研究所
神経細胞の突起が伸びる方向を指示する物質を同定
-幅約10μm領域でのイノシトール三リン酸(IP3)の濃度こう配が鍵-
ポイント
- 微細な神経突起先端部の、IP3の濃度こう配の検出に成功
- IP3の濃度こう配が神経突起の伸びる方向を変えることを発見
- 多種多彩な細胞の移動メカニズムにかかわる普遍的な概念を提供
要旨
独立行政法人理化学研究所(野依良治理事長)は、神経細胞の微細領域(幅約10μm)でイノシトール三リン酸(IP3)※1の濃度のこう配を検出することに成功し、この濃度こう配が神経突起の伸びる方向を変えていることを、世界で初めて発見しました。理研脳科学総合研究センター(利根川進センター長)神経成長機構研究チームの上口裕之チームリーダーと秋山博紀リサーチアソシエイトが、発生神経生物研究チーム(御子柴克彦チームリーダー)と共同で研究した成果です。
運動をはじめ五感、記憶、学習などをつかさどる脳神経系の中心として機能する神経回路は、多数の神経細胞から伸びた神経突起※2が精巧に絡み合って構築されます。この過程で、成長円錐※3と呼ばれる神経突起の先端部が、その細胞外の環境に存在する標識を感受し、正しい方向へ移動します。R・レーヴィ・モンタルチーニ博士らが発見した神経成長因子(1986年ノーベル生理学・医学賞)は、成長円錐を引き寄せる標識として働くことが確認された最初の分子です。成長円錐が周囲にある神経成長因子の濃度差を検知すると、神経突起は神経成長因子の濃度が高い方向へ伸びていきます。しかし、この方向転換を担う分子メカニズムは長年の謎でした。
研究チームは、IP3感受性タンパク質を成長円錐に組み込み、この成長円錐の片側から多くの神経成長因子を投与する実験を行い、細胞外の神経成長因子の濃度こう配が、細胞内IP3の濃度こう配を作り出す様子を見いだしました。これは、成長円錐のような微細な領域では、IP3濃度はほぼ均一であるという従来の仮説を覆す発見です。さらに、成長円錐の内部にIP3の濃度こう配を人工的に作製すると、神経成長因子が存在しない状況であっても、成長円錐はIP3濃度が高い側へ回りこむことを見いだしました。
今回の発見は、神経突起を正しい方向へ誘導するためのシグナル伝達を明らかにするとともに、神経回路の構築機構の解明や損傷した神経回路を修復するための技術開発への貢献が期待される成果です。さらに、免疫系や腫瘍などの非神経細胞が移動するためのメカニズム解明への概念を提供します。
本成果は、米国の科学雑誌『Science Signaling(サイエンス・シグナリング)』のオンライン版(7月14日号:日本時間7月15日)に掲載されます。
背景
脳神経系の中心を担う神経回路は、多数の神経細胞から伸びた神経突起が絡み合った連絡網ですが、この連絡網を正確に構築する仕組み、特に神経突起を目的の方向へ誘導するメカニズムは十分に解明されていません。成長円錐と呼ばれる神経細胞の突起の先端部は、アメーバ状に広がった運動性に富む構造体で(図1)、その周囲環境に存在する標識を感受し、目的の方向へ移動します。神経成長因子は、さまざまな細胞から細胞外に放出されるタンパク質で、成長円錐を引き寄せる標識として働くことが確認された最初の分子です。成長円錐の幅は10~20μm程度ですが、成長円錐の両側で濃度が異なる神経成長因子に遭遇すると、成長円錐は神経成長因子の濃度が高い側へ回りこみます。成長円錐は神経成長因子のわずか1%未満の濃度差を検出し、正しい移動方向を選択することができます。このためには、細胞外にある神経成長因子などの濃度こう配を、成長円錐内部にある何らかのシグナル伝達分子の濃度こう配に変換する必要がありますが(図2)、この仕組みは明らかにされていませんでした。
神経成長因子は、成長円錐の表面にあるTrkA※4と呼ばれるタンパク質に結合します。神経成長因子が結合したTrkAは、細胞内に多種多彩なシグナルを伝えますが、その1つとして、イノシトール三リン酸(IP3)を産生する酵素の働きを高めます。このことから、細胞外の神経成長因子の濃度こう配は、細胞内でのIP3の産生速度に影響することが示唆されます。しかし、細胞内でのIP3の拡散速度は非常に早いため(283平方μm/秒)、成長円錐のような小さな領域ではIP3濃度のこう配は保持されない(IP3濃度は直ちに均一になる)と推測されていました。研究チームは、この従来の仮説に挑戦し、IP3濃度のこう配が成長円錐の回りこむ方向を決定していることの証明に取り組みました。
研究手法と成果
研究チームは、共同研究者である発生神経生物研究チームの御子柴克彦チームリーダーと松浦徹研究員らが開発した、IP3感受性蛍光タンパク質(IRIS-1)の遺伝子をニワトリ脊髄感覚神経節の神経細胞に導入し、この神経細胞から伸びた成長円錐のIRIS-1を測定しました。IRIS-1には、2種類の蛍光タンパク質が連結されていて、IP3濃度に応じてこれら蛍光タンパク質の間の蛍光共鳴エネルギー移動(FRET)※5の効率が変化します。顕微鏡で撮影した成長円錐の画像の各画素(ピクセル)からFRET効率を算出し、生きた成長円錐でのIP3濃度の分布をリアルタイムで可視化することに成功しました。次に、成長円錐の片側に、より多くの神経成長因子を投与しながらFRET効率の変動を追跡したところ、神経成長因子の濃度こう配に応じて細胞内IP3の濃度こう配が作り出されていました。より高濃度の神経成長因子に接した側で、より高濃度のIP3を検出しました(図3)。
逆に、神経細胞を薬剤で処理してIP3の産生あるいはIP3の働きを阻害すると、神経突起は神経成長因子の濃度こう配に反応せずにまっすぐ伸びていきました(図4)。これにより、神経成長因子はIP3の濃度こう配を介して成長円錐を引き寄せることが判明しました。
次に、成長円錐でのIP3濃度こう配の形成が、神経突起の伸びる方向を転換するための十分条件であるか否かを検証しました。まず、紫外線の照射を受けるとIP3を放出する化合物(ケージドIP3)を成長円錐の内部に注入します。次に成長円錐の片側に紫外線レーザーを照射して人工的にIP3を生成し、レーザーを照射しない反対側とのIP3の濃度差を作製しました。すると、このIP3濃度こう配に応答して、成長円錐はIP3濃度が高い側へ向かって回りこみました(図5)。この実験で、IP3濃度こう配は成長円錐の回りこみを誘発することを明らかにしました。
さらに研究チームは、成長円錐で作り出されたIP3濃度のこう配がカルシウムイオン(Ca2+)濃度のこう配へと変換し、このCa2+濃度こう配が、成長円錐の回りこみを駆動することを発見しました。Ca2+濃度こう配により、成長円錐が回りこむメカニズムに関しては、研究チームの成果としてすでに公表しています(2006年12月11日プレスリリース)。今回の研究により、神経成長因子が神経突起を引き寄せるための、細胞内シグナル伝達の全体像を明らかにすることができました。
今後の期待
神経突起の伸びる方向を制御する分子メカニズムの発見は、発生期に神経回路が作られる仕組みを理解し、また神経突起を誘導して損傷した神経回路を再構築するための技術開発に役立つ大変意義のある成果です。さらに、神経回路の形成異常を伴う先天性脳奇形の病因解明や、神経回路の破たんの関与が示唆されている自閉症や統合失調症などの精神疾患の病態解明にも貢献することが期待されます。
神経突起に限らず生体内のさまざまな細胞は、特定の方向へ移動する能力を備えており、この細胞移動の能力は、各種臓器の発生、免疫、創傷治癒などの生理機能に必須であるとともに、炎症やがん転移などの病態にも関与します。IP3 の濃度こう配が細胞移動の方向を決定するという基本原理は、今後、多種多彩な細胞の移動メカニズムを研究する上で重要な概念を提供します。
発表者
理化学研究所
脳科学総合研究センター 神経成長機構研究チーム
チームリーダー 上口 裕之(かみぐち ひろゆき)
Tel: 048-467-6137 / Fax: 048-467-9795
お問い合わせ先
脳科学研究推進部 納富 さより(のうどみ さより)Tel: 048-467-9757 / Fax: 048-462-4914
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.イノシトール三リン酸(IP3)
ホルモンや神経伝達物質などが細胞の表面に結合すると、細胞膜を構成するリン脂質が分解されて細胞内でIP3が産生される。IP3は代表的な細胞内シグナル伝達分子であり、Ca2+貯蔵庫から細胞質へのCa2+の放出を促進する。 - 2.神経突起
神経細胞から伸びる細長い突起で、電気的情報を遠隔部位に存在するほかの神経細胞や筋肉細胞などに伝える。神経突起の連絡網(神経回路)が、脳神経系の働きの中心的役割を担っている。 - 3.成長円錐
伸長している神経突起の先端部に形成される、手のひらのような形をした構造体。細胞外に存在するさまざまな標識分子を感知し、正しい道のりを移動することにより、神経突起を目的の方向へ誘導する。 - 4.TrkA
神経成長因子の受容体として機能する細胞膜貫通タンパク質。神経成長因子が結合したTrkAは、IP3を産生する酵素の働きを高めるなど、さまざまなシグナルを細胞内に伝達する。 - 5.蛍光共鳴エネルギー移動(FRET)
近接した2個の色素分子の間でエネルギーが直接移動する現象。生物学分野では、シアン蛍光タンパク質と黄色蛍光タンパク質の間のFRETを観測することで、分子内/分子間相互作用の検出に応用されることが多い。
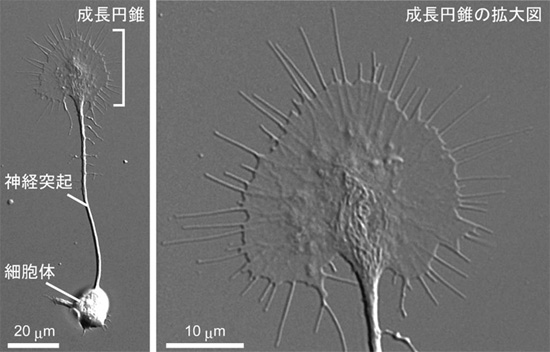
図1 神経突起と成長円錐の形態

図2 分子濃度こう配の役割を示す概念図
図上方へ向かって伸びている神経突起が神経成長因子の濃度こう配(青色部分)に遭遇すると、成長円錐の内部には、赤で示すようなシグナル伝達分子の濃度こう配が作られる。その結果、成長円錐は右方向へ回りこみ、神経突起は神経成長因子の濃度がより高い方向へ伸びていく。
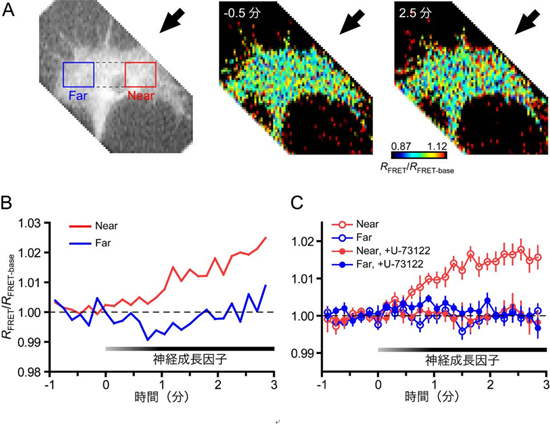
図3 成長円錐でのIP3濃度分布の時間変化
(A)成長円錐にIRIS-1(IP3感受性蛍光タンパク質)を導入し、その右側(矢印)から神経成長因子を投与した。カラー写真はIRIS-1のFRET効率(IP3濃度の指標)の擬似カラー表示であり、神経成長因子の濃度こう配を作製する0.5分前および2.5分後の成長円錐を示す。赤色に近づくに従い、IP3濃度が上昇したことを表す。左端の画像上の赤線と青線は、(B)でFRET効率の平均値を算出するために用いた領域を示す。
(B)神経成長因子の濃度こう配を作製する前後のIP3濃度の時間変化。0分の時点で神経成長因子の投与を開始した。グラフの縦軸はIRIS-1のFRET効率(IP3濃度の指標)を表す。グラフの赤線と青線はそれぞれ、(A)の赤線(Near)と青線(Far)で囲まれた領域でのFRET効率の平均値を示す。より高濃度の神経成長因子に接する赤線領域ではIP3濃度は上昇したが、反対側の青線領域でのIP3濃度はほぼ一定であった。
(C)同様の条件で、複数の成長円錐のデータを測定した。IP3の産生を阻害する薬剤(U-73122)で処理した成長円錐では、神経成長因子を投与してもIP3濃度は変化しなかった。
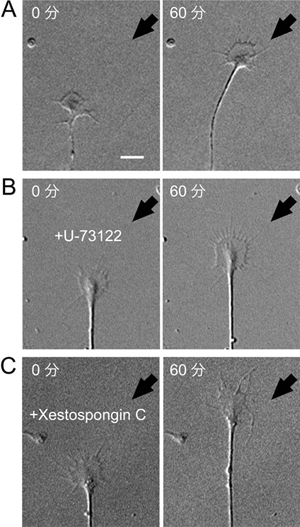
図4 IP3は成長円錐の回りこみに必要
(A)神経成長因子の濃度こう配(矢印側が高濃度)が神経突起を引き寄せる様子を示す。各写真の左上に、神経成長因子の投与開始後の時間(分)を記す。スケールバーは10μmに相当。
(B)IP3の産生を阻害する薬剤(U-73122)で処理した神経突起は、神経成長因子の濃度こう配(矢印側が高濃度)に反応せず直進した。
(C)IP3の働きを阻害する薬剤(Xestospongin C)で処理した神経突起は、神経成長因子の濃度こう配(矢印側が高濃度)に反応せず直進した。
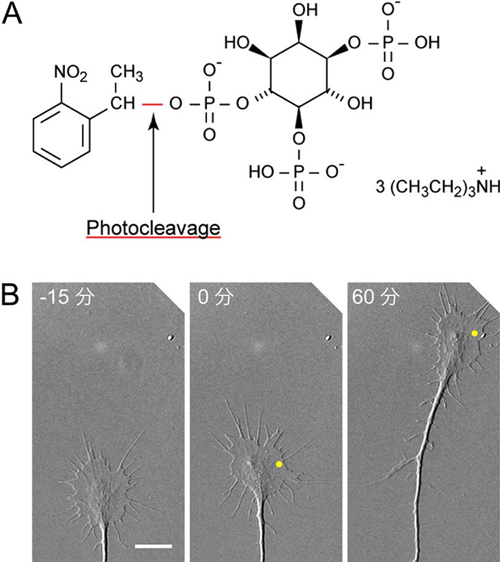
図5 IP3濃度こう配は成長円錐の回りこみを誘発する
(A)ケージドIP3の化学構造。ケージドIP3はIP3としての生理活性を持たないが、紫外線照射により赤線で示した結合が切断され、IP3(図右側の分子)が放出される。ケージドIP3を注入した細胞に紫外線を照射することで、細胞の任意の領域でIP3を生成することができる。
(B)ケージドIP3を注入した成長円錐の右側(黄色の点)に紫外線レーザーを照射してIP3を生成すると、神経突起は右方向へ伸長した。各写真の左上は、紫外線レーザー照射を開始してからの時間(分)。スケールバーは10μmに相当。