要旨
理化学研究所(理研)倉谷形態進化研究室の倉谷滋主任研究員、兵庫医科大学教養部門生物学の菅原文昭講師(理研倉谷形態進化研究室客員研究員)らの共同研究グループ※は、顎(あご)を持たない脊椎動物「円口類」に属するヌタウナギ[1]とヤツメウナギ[2]の脳の発生過程を観察し、これらの動物では見つかっていなかった脳の中の2領域を新たに発見しました。これにより、段階的に進化してきたと考えられてきた脳の各領域のほとんどが、5億年以上前にすでに成立していたことを明らかにしました。
脳は細かく領域化された複雑な器官ですが、各領域が進化の過程でいつ獲得されたのかについては、未解明な点が多く残されています。現在、地球上に生息する脊椎動物の中で最初に分岐したのは、顎を持たない「円口類」と呼ばれる動物群です。円口類と、ヒトのように顎を持つ「顎口(がっこう)類」との比較により、脊椎動物の脳の初期進化を解明できると考えられます。
これまでの研究から、円口類の一種ヤツメウナギには大脳基底核[3]の一部である内側基底核隆起[4]が見られず、小脳[5]は未発達であることが分かっていました。そのため、2つの領域は、顎口類になってから新しく発達したと考えられていました。あるいは、これらの領域がヤツメウナギの進化の過程で二次的に退化した可能性もあります。それを調べるには、円口類のもう1つの系統、ヌタウナギの脳との比較が重要と考えられました。しかし、ヌタウナギの胚を得ることは非常に難しいため、近年まで脳の発生過程についての研究は全く進んでいませんでした。
そこで共同研究グループはまず、実験室内で得ることに成功した希少なヌタウナギ胚を使用し、ヌタウナギの脳の発生過程を観察しました。特に、脳の各領域を特徴付ける遺伝子の発現と、脳内の神経線維の走行の解析を行いました。その結果、ヤツメウナギで見つかっていなかった内側基底核隆起がヌタウナギの胚には存在することが分かりました。さらに、菱脳唇(りょうのうしん)と呼ばれる小脳が発生する場となる部分と、小脳を構成するニューロンが発生するために不可欠な遺伝子の発現も見つかりました。これらの結果を踏まえ、ヤツメウナギ胚を再解析したところ、これまで存在しないとされていた、これら2つの領域(内側基底核隆起と菱脳唇)がヤツメウナギにも存在することが明らかになりました。
以上から、これまで段階的に進化してきたと考えられてきた脊椎動物の脳の各領域の多くが、5億年以上前という脊椎動物の進化過程の極めて早い段階に、すでに成立していたことが明らかになりました。
本研究は、国際科学雑誌『Nature』に掲載されるのに先立ち、オンライン版(2月15日付け:日本時間2月16日)に掲載されます。
※共同研究グループ
理化学研究所
倉谷形態進化研究室
主任研究員 倉谷 滋(くらたに しげる)
研究員 フアン・パスクァル・アナヤ(Juan Pascual-Anaya)
テクニカルスタッフ(研究当時)大石 康博(おおいし やすひろ)(現マックスプランクフロリダ研究所)
研究員(研究当時)青田 伸一(あおた しんいち)
研究員(研究当時) 足立 礼孝 (あだち のりたか)(現シカゴ大学)
訪問研究員 高木 亙(たかぎ わたる)
テクニカルスタッフⅠ 平井 珠美(ひらい たまみ)
ライフサイエンス技術基盤研究センター 分子配列比較解析ユニット
ユニットリーダー 工樂 樹洋(くらく しげひろ)
兵庫医科大学 教養部門生物学
講師 菅原 文昭(すがはら ふみあき)(理研倉谷形態進化研究室 客員研究員)
新潟大学大学院 医歯学総合研究科
教授 佐藤 昇(さとう のぼる)
愛媛大学大学院 理工学研究科
准教授 村上 安則(むらかみ やすのり)
背景
脊椎動物の脳は、高度に領域化された非常に複雑な器官ですが、その成立や各領域の進化の過程には未解明な点が多く残されています。脳は柔らかいため化石として残りにくく、絶滅した初期の脊椎動物の化石から分かることは限られるためです。一方、現在も地球上に生息している脊椎動物は、ヒトも含めた顎(あご)を持つ「顎口(がっこう)類」と顎を持たない「円口類」に大別でき、これらは5億年以上前に分岐しました。従って、円口類と顎口類の発生過程を比較すれば、脳の進化的変遷を解明できると考えられます。
現生の円口類にはヌタウナギとヤツメウナギの2群がいますが(図1)、これまでの円口類の発生学研究では、胚を得ることが容易なヤツメウナギが使用されてきました。その研究から、円口類と顎口類の脳における最も大きな違いは、以下の2点であると言われていました。
① 円口類は、大脳基底核の最も腹側の領域(内側基底核隆起)を持たない。
② 円口類は、発達した小脳を持たない。
これまで、内側基底核隆起と小脳は、円口類と顎口類が分岐したのち、顎口類の進化の過程で新たに獲得・発達してきたと考えられてきました。しかし、ヤツメウナギの系統で2つの領域が二次的に退化した可能性も否定できません。さらに近年、内側基底核隆起から派生する脳の領域とニューロン種が、ヤツメウナギの成体に見られるという報告もされています。
この問題を解決するためには、円口類に属するもう1つのグループ、ヌタウナギの脳の発生を知る必要があります。しかし、近年までヌタウナギの発生過程は謎に包まれていました。それはヌタウナギが深海に生息し、胚を得るために必要な受精卵の入手が極めて難しかったためです。しかし、共同研究グループは2007年、世界で初めてヌタウナギの人工繁殖に成功し、以来多くの知見を得ることができるようになりました注)。
注)理研CDB科学ニュース
2007年3月19日号 人工飼育環境下でヌタウナギの発生に成功
2011年7月1日号 ヌタウナギに背骨?
2013年1月31日号 ヌタウナギに見る脊椎動物の進化
研究手法と成果
共同研究グループは、島根県江津市で漁を営む柿谷紀(かきたに おさむ)氏の協力により、2011年に島根県浜田市沖で獲った繁殖期のヌタウナギから受精卵の採取に成功しました。それらを研究室内で発生させ、いくつかの胚を手に入れました。そのうち異なる発生段階の2つの胚を使用して組織切片を作製し、in situハイブリダイゼーション[6]による遺伝子の発現パターンと免疫染色[7]による脳の神経線維の走行を調べました。特に脳の各領域を特徴付ける遺伝子の発現と神経線維の走行を解析し、ヌタウナギ胚における脳の詳しい発生地図(発生初期に決まる脳形成のための地図)を初めて作成しました。
その結果、これまで円口類では見つかっていなかった大脳基底核の内側基底核隆起に相当する遺伝子(Nkx2.1、ヘッジホッグ)の発現が観察され、この領域がヌタウナギ胚に存在することが明らかになりました(図2)。さらに、菱脳唇と呼ばれる小脳が発生する場に特徴的な遺伝子Pax6の発現と、小脳の層構造の1つを構成する顆粒細胞の分化に必須である転写因子Atoh1遺伝子の発現も同時に見られました(図2)。こうして、ヌタウナギに、小脳と同等の領域と遺伝子発現が存在することが明らかになりました。
この結果を踏まえ、共同研究グループはヤツメウナギ胚を詳しく再解析しました。近年公開されたヤツメウナギのゲノム情報を利用して、これまで知られていなかった多くの遺伝子を発見し、その発現パターンを網羅的に解析しました。その結果、これまでの研究では見つからなかった大脳基底核の内側基底核隆起における遺伝子発現(Nkx2.1)と、菱脳唇における遺伝子発現(Pax6、Wnt1)および、小脳の層を構成するニューロンの分化に必須の遺伝子発現(Atoh1:顆粒細胞、Ptf1a:プルキンエ細胞)の全てが、ヤツメウナギにもあることが分かりました(図2)。
以上の結果から、これまで円口類の分岐以降に獲得したと考えられていた内側基底核隆起と菱脳唇が、円口類の両グループに存在することが分かりました。これらの脳領域の獲得は、円口類と顎口類の共通祖先が生きていた頃、すなわち5億年以上前にまでさかのぼることが明らかになりました。これは、脊椎動物の脳が段階的に複雑化してきたという従来の理解とは異なり、脊椎動物の成立初期にその基本パターンの大部分が成立していたという、進化の新たなシナリオを示しています(図3)。
今後の期待
今回、大脳基底核の領域と、小脳獲得の前段階ともいえる領域とニューロンの分化に必要な遺伝子発現が、円口類のヌタウナギとヤツメウナギの両系統に存在することが分かりました。これにより、これらの領域は脊椎動物の進化の極めて初期に獲得されたことが示されました。これは「ヒトに至る過程で脳はどう進化したのか」という大きな謎に迫る成果です。今後の課題として、例えば、大脳新皮質の起源が上げられます。大脳新皮質はほ乳類で特に発達した領域ですが、は虫類や鳥類でも類似した領域が見つかっています。この大脳新皮質の起源はどのような進化的変遷を経て、ヒトの「知性」がどのように生まれたのか、非常に興味のあるテーマです。
また、全ての顎口類に見られ、体性感覚や平衡感覚を受容し運動を制御するための発達した層構造を持つ小脳が、どのような発生プログラムの変遷を経て獲得されたのか、解明されていません。今後、小脳獲得の「前夜」の状態をいまだに保持していることが分かった円口類と小脳を持つ動物の中で最も「古い」系統であるといわれる軟骨魚類(サメやエイなど)とを比較することで、詳細な進化の経緯の解明が期待できます。
原論文情報
- Fumiaki Sugahara, Juan Pascual-Anaya, Yasuhiro Oisi, Shigehiro Kuraku, Shin-ichi Aota, Noritaka Adachi, Wataru Takagi, Tamami Hirai, Noboru Sato, Yasunori Murakami & Shigeru Kuratani, "Evidence from cyclostomes for complex regionalization of the ancestral vertebrate brain", Nature, doi: 10.1038/nature16518
発表者
理化学研究所
主任研究員研究室 倉谷形態進化研究室
主任研究員 倉谷 滋(くらたに しげる)
兵庫医科大学 教養部門生物学
講師 菅原 文昭(すがはら ふみあき)
 共同研究グループのメンバー(一部)
共同研究グループのメンバー(一部)左から平井珠美、高木亙、工樂樹洋、倉谷滋、菅原文昭、フアン・パスクァル・アナヤ
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
兵庫医科大学 経営企画室 広報課
Tel: 0798-45-6655 / Fax: 0798-45-6219
kouhou[at]hyo-med.ac.jp(※[at]は@に置き換えてください。)
補足説明
- 1.ヌタウナギ
円口類に属する顎のない脊椎動物の一群。細長い体型のため「ウナギ」と呼ばれるが、真骨魚ウナギ目の仲間ではない。脊椎骨が退化しているため、かつては脊椎動物の前段階の動物と見なされていた。ほとんどの種が深海に生息する。本共同研究グループは、日本産ヌタウナギ( Eptatretus burgeri)から世界で唯一、実験室内での胚の取得に成功している。眼や松果体(内分泌器)は退化しており、粘液腺から粘液を放出し、捕食や防御に用いる。 - 2.ヤツメウナギ
ヌタウナギと同じく、円口類に属する一群。吸盤状の口を持ち、眼の後ろに開口する7対のえら孔が眼になぞらえられるため、この名がある。本研究では日本産カワヤツメウナギ( Lethenteron japonicum)を使用した。 - 3.大脳基底核
大脳皮質と視床、脳幹を結び付けている神経核の集まり。主に運動の調節に関わる。 - 4.内側基底核隆起
大脳基底核の一部位の発生原基の名称で、MGE(Medial Ganglionic Eminence )とも呼ばれる。成体では淡蒼球(たんそうきゅう)に分化し、運動パターンの選択を行う。さらに、大脳皮質に分布する抑制ニューロンの発生源となる。 - 5.小脳
体性感覚や平衡感覚を受容し、運動を制御する脳の部位。後脳の背側(菱脳唇)の前方より発生し、顆粒層、プルキンエ細胞層、分子層の3層からなる。このうち、顆粒細胞とプルキンエ細胞の分化には転写因子 Atoh1、 Ptf1aがそれぞれ重要な役割を果たすことが知られている。全ての顎口類に小脳が存在する一方、円口類では層をなす小脳は見られず、小脳交連など一部の神経線維が認められるに留まる。 - 6.in situハイブリダイゼーション
特定の遺伝子が、体の中のどこで発現しているかを観察する方法。 - 7.免疫染色
特定のタンパク質が、体の中のどこで発現しているのかを観察する手法。抗体の持つ特性を利用するため、免疫染色と呼ばれる。

図1 ヌタウナギ(奥)とヤツメウナギ(手前)
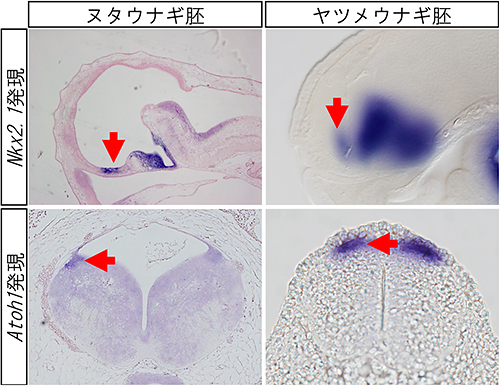
図2 ヌタウナギとヤツメウナギの遺伝子発現パターン
上段:大脳の内側基底核隆起を特徴づけるNkx2.1遺伝子の発現(赤色の矢印)。
下段:小脳が発生する場の菱脳唇を特徴付けるAtoh1遺伝子の発現(赤色の矢印)。
これら遺伝子の発現から、円口類(ヌタウナギ、ヤツメウナギ)の両系統に内側基底核隆起と菱脳唇が存在することが明らかとなった。
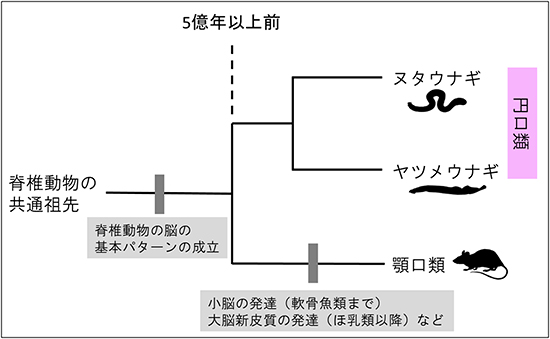
図3 脊椎動物における脳の進化のシナリオ
脊椎動物の脳の領域化は、円口類と顎口類が分岐する以前にその多くが成立していた。特に本研究では、内側基底核隆起における遺伝子発現と、菱脳唇における遺伝子発現および小脳を構成するニューロンの分化に必須の遺伝子発現の全てが、円口類との分岐以前に獲得されていたことが示された。円口類との分岐以降、層構造を持った真の小脳が軟骨魚類(サメやエイなど)の分岐までに獲得され、さらにほ乳類の進化の過程で大脳新皮質が著しく発達した。