要旨
理化学研究所(理研)グローバル研究クラスタ糖鎖構造生物学研究チームの山口芳樹チームリーダー、長江雅倫研究員らの共同研究グループ※は、形に揺らぎのある糖鎖を特定のタンパク質「レクチン[1]」と結合させて固定することにより、折れ曲がった状態の糖鎖構造を原子レベルで可視化することに成功しました。
糖鎖は生体内で、細胞同士や細胞の外から内への情報伝達、タンパク質の機能調節など多くの重要な役割を果たしています。糖鎖は一般に柔軟な構造をしており、1つの糖鎖がいくつもの形をとることが知られています。つまり、形に揺らぎがあるのです。しかし、糖鎖の形は非常に速い速度で相互に変換しており、糖鎖の個々の形を実験的に正確に捉えることはこれまで困難でした。
共同研究グループは、揺らぎのある糖鎖を特定のレクチンと結合させることにより、揺らぎを止めた状態の糖鎖の構造を可視化しようと考えました。糖鎖のモデルとして、これまで折れ曲がった構造をとると予想されていた「バイセクト型糖鎖[2]」を選びました。レクチンは、バイセクト型糖鎖に結合する互いに無関係な2種類、「Calsepaレクチン」と「E4-PHAレクチン」を使用しました。バイセクト型糖鎖を各レクチンとの複合体を作製して、X線結晶構造解析[3]およびNMR(核磁気共鳴)解析[4]を行い、構造を調べました。その結果、どちらのバイセクト型糖鎖-レクチン複合体も、糖鎖は折れ曲がった構造をしていることが分かり、原子レベルでの可視化に成功しました。
本研究は、糖鎖の形の揺らぎと機能を理解するための重要な知見になるとともに、糖鎖とタンパク質の相互作用原理や、糖鎖がタンパク質を調節するメカニズムの理解につながると期待できます。
成果は、英国のオンライン科学雑誌『Scientific Reports』(3月14日付け:日本時間3月14日)に掲載されました。
※共同研究グループ
理化学研究所 グローバル研究クラスタ
糖鎖構造生物学研究チーム
チームリーダー 山口 芳樹(やまぐち よしき)
研究員 長江 雅倫(ながえ まさみち)
研究員 金川 真由美(かながわ まゆみ)
人材派遣 森田 加奈(もりた かな)
疾患糖鎖研究チーム
特別研究員 木塚 康彦(きづか やすひこ)
チームリーダー 谷口 直之(たにぐち なおゆき)
大阪大学大学院 理学研究科
講師 花島 慎弥(はなしま しんや)
背景
糖鎖は糖(単糖)がつながった生体分子であり、主に細胞の表面に存在しています。その多くは細胞膜に埋め込まれたタンパク質や脂質に結合し、細胞同士や細胞の外から内への情報伝達、タンパク質の品質管理・機能調節など、生体内でさまざまな重要な役割を果たしています。糖鎖は柔軟な構造をとることが知られており、通常は水溶液中で複数の安定な形をとっています。つまり、形に揺らぎがあるのです。揺らぎも含めて糖鎖の形を捉えることは、糖鎖の機能を理解する上で極めて重要です。
しかし、糖鎖の形の相互変換の速度はとても速く、個々の形を実験的に捉えることは困難でした。特に存在する確率が低い形を捉えることは、検出感度や分解能などの制限から非常に困難でした。
研究手法と成果
共同研究グループは、揺らぎのある糖鎖を特定のレクチン(糖鎖を特異的に認識し結合するタンパク質の総称)と結合させることにより、揺らぎを止めた状態の糖鎖の構造を可視化しようと考えました。糖鎖は、これまで折れ曲がった構造をとると予想されていた「バイセクト型糖鎖」をモデル糖鎖として選びました。レクチンは、互いに無関係な「Calsepaレクチン」と「E4-PHAレクチン」の2種を使用しました。
まず、化学合成と天然糖タンパク質からの分離精製によりバイセクト型糖鎖を調製しました。その後、バイセクト型糖鎖と各レクチンとの複合体を作製して、X線結晶構造解析を行いました。その結果、どちらのレクチン-糖鎖複合体においても、糖鎖の2本の枝のうちの1本の「1-6アーム」と「1-3アーム」が反対の向きを向いた、折れ曲がり構造をしていることが分かりました(図1)。これまでの研究で、分岐型の糖鎖がとり得る形は、主に伸長型と折れ曲がり型の2つが存在すると予測されていました。伸長型はすでに立体構造が報告されていますが、折れ曲がり型はその存在確率が低いため、原子レベルで立体構造を解析した報告はありませんでした。
続いて、糖鎖の折れ曲がり構造が水溶液中で存在することを確認するために溶液NMR法を適用しました。バイセクト型糖鎖のユニットとCalsepaレクチンを混合し、転移NOE法[5]によってCalsepaレクチンに結合した状態のバイセクト型糖鎖の立体構造を調べました。その結果、レクチン非存在下では観測されないようなNOEの信号が、Calsepaレクチン存在下では観測されました(図2)。このNOEの信号は折れ曲がり構造においてのみ観測される信号です。このことから、水溶液中においてもバイセクト型糖鎖はCalsepaレクチンとの結合時に折れ曲がり構造をとることが確認されました。
今後の期待
本成果は、形の揺らぎが糖鎖の機能に果たす役割を理解するための重要な知見になります。また、糖鎖とタンパク質の相互作用原理や糖鎖がタンパク質の機能を調節するメカニズムの理解につながり、将来的には糖タンパク質を利用した医薬品の開発に貢献すると期待できます。
原論文情報
- Masamichi Nagae, Mayumi Kanagawa, Kana Morita-Matsumoto, Shinya Hanashima, Yasuhiko Kizuka, Naoyuki Taniguchi and Yoshiki Yamaguchi, "Atomic visualization of a flipped-back conformation of bisected glycans bound to specific lectins", Scientific Reports, doi: 10.1038/srep22973
発表者
理化学研究所
グローバル研究クラスタ 理研-マックスプランク連携研究センター システム糖鎖生物学研究グループ 糖鎖構造生物学研究チーム
チームリーダー 山口 芳樹(やまぐち よしき)
研究員 長江 雅倫(ながえ まさみち)
 山口 芳樹
山口 芳樹
 長江 雅倫
長江 雅倫
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.レクチン
糖と結合するタンパク質のこと。Calsepa、E4-PHAもレクチンの1種である。 - 2.バイセクト型糖鎖
タンパク質のアミノ酸、アスパラギンの側鎖に結合する分岐型糖鎖の1つ。2本の枝の分岐部分のマンノースに N-アセチルグルコサミンが結合した糖鎖のこと。 - 3.X線結晶構造解析
結晶内の分子構造を解明する手法。1Å程度の波長のX線を使い、タンパク質など物質が規則正しく並んだ結晶によって回折されるX線回折データを得て、回折されたX線の強度を詳しく解析し結晶構造を知る方法。 - 4.NMR(核磁気共鳴)解析
静磁場に置かれた原子核の共鳴を観測し、分子の構造や運動状態などの性質を調べる分光法。溶媒に分子を溶解させて計測する溶液NMR法や固体状態の分子を計測する固体NMR法などがあり、幅広い状態の試料を計測することができる。 - 5.転移NOE法
NOEは、核オーバーハウザー効果(Nuclear Overhauser Effect)の略。空間的に近接した水素原子(通常5Å以内)のペアで観測されるNMR現象のこと。NOEの信号強度が水素原子間の距離に依存することから、NOEは対象分子の立体構造を決定する際の重要なパラメータとなっている。転移NOE法は、タンパク質などの高分子に結合した低分子リガンドの立体構造を調べる方法。本研究では、レクチンに結合した状態の糖鎖において観測されるNOEを用いて解析した。
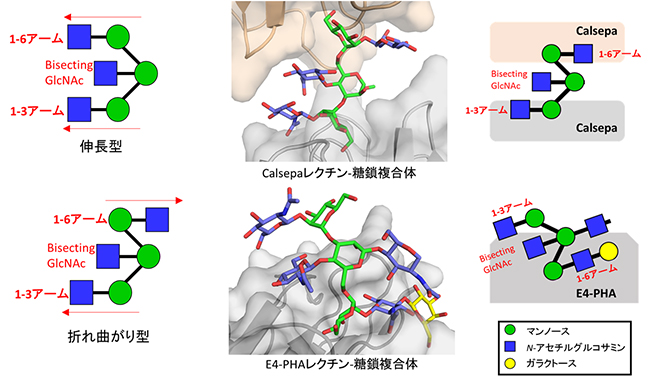
図1 バイセクト型糖鎖とCalsepaレクチンおよびE4-PHAレクチンの複合体の結晶構造
左:バイセクト型糖鎖のモデル。2本の枝の分岐部分のマンノースにN-アセチルグルコサミン(bisecting GlcNAc)が結合している。伸長型(上)と折れ曲がり型(下)の両方の形をとる。伸長型では、2本の枝うちの1本の枝の1-6アームと1-3アームが赤い矢印で示すように、同じ向きを向いた構造をしている。折れ曲がり型では、1-6アームと1-3アームが赤い矢印で示すように、反対の向きを向いた構造をしている。
中:X線結晶構造解析により分かった、Calsepaレクチン-糖鎖複合体(上)とE4-PHAレクチン-糖鎖複合体(下)の立体構造。
右:中の図の糖鎖の立体構造を模式化した図。どちらの複合体も、糖鎖の1-6アームと1-3アームが反対の向きを向いた折れ曲がり構造をしている。
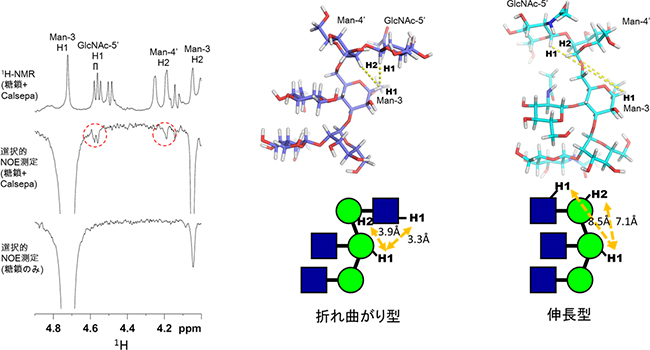
図2 折れ曲がり構造の存在を示すバイセクト型糖鎖NOEシグナルの検出
左:バイセクト型糖鎖ユニットの1H-NMRスペクトル。Calsepaレクチン存在下の糖鎖の1H-NMRスペクトル(上段)、選択的NOE(核オーバーハウザー効果)スペクトル(中段)、糖鎖のみの選択的NOEスペクトル(下段)。中段では、レクチン存在下において、Man-3(マンノース-3)のH1に由来するシグナルを選択的に反転させた。すると2つの赤い点線の丸で示すように、GlcNAc-5’(N-アセチルグルコサミン-5’)のH1およびMan-4’(マンノース-4’)のH2において転移NOEシグナルが観測された。この結果はバイセクト型糖鎖ユニットがレクチンと結合しているときに折れ曲がり構造をとることを意味する。それぞれの水素(H)は、中図の上の分子構造を参照。
中と右:折れ曲がり型と伸長型における分子構造(上段)と、Man-3のH1とGlcNAc-5’のH1間、Man-3のH1とMan-4’のH2間の距離(下段)。折れ曲がり型では、その水素原子間の距離が5オングストローム(Å、1Åは100億分の1メートル)以内と短く(各3.3Å、3.9Å)、NOEが観測される。一方、伸長型では距離が長くなり(5Å以上、各8.5Å、7.1Å)、NOEが観測されない。