要旨
理化学研究所(理研)環境資源科学研究センター機能開発研究グループの中野雄司専任研究員、山上あゆみ研究員、篠崎一雄グループディレクター、東京大学大学院農学生命科学研究科の浅見忠男教授らの共同研究グループは、植物細胞の伸長を促進する新しいタンパク質「BIL4」を発見し、BIL4が植物ステロイドホルモン[1]「ブラシノステロイド[2]」のシグナル伝達を介して植物細胞の伸長を制御する仕組みを明らかにしました。
ブラシノステロイドは、植物の成長を促す上で重要な役割を果しています。しかし、非常に高価なため農業や植物バイオマスの増産に直接利用されていません。また、ブラシノステロイドが植物内でどのような「シグナル伝達」を行っているかはよく分かっておらず、基礎研究と応用研究の両面における解明が求められています。
今回、共同研究グループは、ブラシノステロイドの生合成を自在に制御できる阻害剤「ブラシナゾール(Brz)[3]」を用いたケミカルバイオロジー[4]の手法を使い、ブラシノステロイドのシグナル伝達を活性化し、制御するタンパク質「BIL4」を発見しました。BIL4は7回膜貫通型(細胞膜を7回貫通する構造)を持つ膜タンパク質で、相同性タンパク質[5]がトマトやイネ、コムギなどの植物だけでなく、ヒトやマウスなどの動物にも存在しており、進化的に広く保存されています。BIL4の細胞内での動きを調べたところ、ブラシノステロイド受容体「BRI1」と、細胞内小器官[6]のエンドソーム[7]において相互作用することが分かりました。BRI1は細胞膜に存在し、細胞外からのブラシノステロイドを感知してシグナルを細胞内に伝達する起点となり、植物の成長において非常に重要な因子であると考えられています。また、BIL4はエンドサイトーシス[8]で細胞内に取り込まれた後、液胞[9]に運ばれて分解されるという制御を受けることが近年明らかになっています。以上のことから、BIL4はBRI1の分解を防ぐ機能を持つこと、その分解抑制によってブラシノステロイドのシグナル伝達を活性化すること、さらにシグナル伝達の活性化により植物の胚軸(発芽した幼植物体の茎部分)や緑葉の細胞伸長を促す機能を持つことが明らかになりました。
本成果により、植物バイオマスや有用作物の葉のサイズや草丈を自在に制御する技術の開発が進むと期待できます。
本研究は、英国のオンライン科学雑誌『Scientific Reports』(7月18日付け)に掲載されます。
本研究は、科学技術振興機構(JST)戦略的創造研究推進事業(CREST)「二酸化炭素資源化を目指した植物の物質生産力強化と生産物活用のための基盤技術の創出」研究領域(研究総括:磯貝彰(奈良先端科学技術大学院大学 名誉教授))における研究課題「植物ホルモン間クロストークと化学・生物学的制御技術を利用したバイオマス高生産性植物の開発」(研究代表者:浅見忠男、共同研究者:中野雄司)の一環として行われました。
※共同研究グループ
理化学研究所 環境資源科学研究センター
機能開発研究グループ
専任研究員 中野 雄司(なかの たけし)
研究員 山上 あゆみ(やまがみ あゆみ)
グループディレクター 篠崎 一雄(しのざき かずお)
ケミカルバイオロジー研究グループ
グループディレクター 長田 裕之(おさだ ひろゆき)
合成ゲノミクス研究グループ
グループディレクター 松井 南(まつい みなみ)
理化学研究所 基幹研究所
中野生体膜研究室
主任研究員(研究当時)中野 明彦(なかの あきひこ)(現 光量子工学研究領域 生細胞超解像イメージング研究チーム チームリーダー
現本務 東京大学大学院 理学系研究科 教授)
専任研究員(研究当時)齊藤 知恵子(さいとう ちえこ)(現 科学技術振興機構研究開発戦略センター フェロー)
東京大学大学院
農学生命科学研究科 応用生命化学専攻 生物制御化学研究室
教授 浅見 忠男(あさみ ただお)
東京大学大学院
理学系研究科 生物科学専攻
助教 植村 知博(うえむら ともひろ)
お茶の水女子大学大学院
人間文化創成科学研究科 ライフサイエンス専攻
教授 作田 正明(さくた まさあき)
背景
植物ステロイドホルモンである「ブラシノステロイド」は、植物の成長に重要な役割を果たしています。しかし、ブラシノステロイドは非常に高価で、農業や植物バイオマスの増産に直接利用されていません。また、ブラシノステロイド生合成のメカニズムは大部分が明らかになっていますが、ブラシノステロイドが植物内でどのような「シグナル伝達」を行っているかは分かっておらず、基礎研究と応用研究の両面における解明が求められています。
動物から植物までの広い生物種において、多くの生理活性化合物は受容体(レセプター)と呼ばれるセンサー役のタンパク質に感知され、その刺激が細胞内を伝わっていくシグナル伝達という機構によって、生理活性が発現されます。ブラシノステロイドにおいては、「BRI1」という1回膜貫通型(細胞膜を1回貫通する構造)のリン酸化酵素(キナーゼ)が細胞表面に局在し、細胞外からくるブラシノステロイドを感知する受容体としての役割を持つことが分かっていました。近年、BRI1は細胞膜だけでなく、エンドソームと呼ばれる細胞膜の内側近くにある細胞小器官にも局在していることが分かっており、局在の生物学的な意味について現在、多くの研究が試みられています。
研究手法と成果
共同研究グループはまず、ブラシノステロイドの生合成を自在に制御できる阻害剤「ブラシナゾール(Brz)」を用い、ケミカルバイオロジーによる実験を行いました。シロイヌナズナの種子をBrz処理条件下で暗所発芽させた野生型株は、通常、短い胚軸(発芽した幼植物体の茎部分)を示します。これに対して、シロイヌナズナ変異体種子群の中から、Brz処理条件下の暗所発芽時に長い胚軸伸長を示す変異体「bil4(Brz-insensitive-long hypocotyl4)」を単離しました。このbil4変異体の遺伝子発現などを調べたところ、bil4変異体では、ブラシノステロイドのシグナル伝達が活性化されていると予測されました(図1)。
bil4変異体の原因遺伝子を探索したところ、新しいタンパク質「BIL4」の高発現が原因であることが分かりました。BIL4は7回膜貫通型(細胞膜を7回貫通する構造)を持つ膜タンパク質で、相同性タンパク質がトマトやイネ、コムギなどの植物に存在するだけでなく、ヒトやマウスなどの動物にも存在し、進化的に広く保存されています。しかし、植物における詳しい機能についてはよく分かっていませんでした。
そこでBIL4の機能を調べるために、BIL4が野生型よりも高発現している変異体(BIL4活性化型変異体)と遺伝子組換え植物(BIL4活性化型形質転換体)を作製し観察したところ、草丈が野生型よりも長くなる傾向がみられました(図2)。また、BIL4が野生型よりも低発現している遺伝子組換え植物(BIL4低発現型形質転換体)は、植物の草丈が野生型よりも短くなる特徴を示しました(図3)。
さらに、BIL4の高発現化もしくは低発現化による植物形態の変化が、細胞レベルではどのような影響が現れるかを調べました。その結果、Brz存在下の暗所発芽条件において、BIL4が野生型よりも高発現している変異体(BIL4活性化型変異)と遺伝子組換え植物(BIL4活性化型形質転換体)の胚軸の表皮細胞は、野生型よりも長いことが分かりました(図4)。
続いて、ロゼッタ葉(成熟した緑葉)の表皮細胞を観察したところ、BIL4が野生型よりも高発現している遺伝子組換え植物(BIL4活性化型形質転換体)の細胞サイズは野生型よりも大きく、BIL41が野生型よりも低発現している遺伝子組換え植物(BIL4低発現型形質転換体)は、細胞サイズが野生型よりも小さいことが分かりました(図5)。これらの結果は、BIL4が胚軸や葉の細胞の伸長を促す活性を持つことを示しています。
次に、BIL4と緑色蛍光タンパク質(GFP)との融合タンパク質(BIL4-GFP)を使って、BIL4の植物細胞内での状態を調べたところ、BIL4は主に細胞質でドット状(点状)の明るい蛍光シグナルとして観察されました。このドット状のシグナルを細胞膜の蛍光染色試薬や植物オルガネラ局在タンパク質マーカーなどを用いて観察した結果、細胞内小器官のエンドソームであることが分かりました。
続いて、BIL4-GFPと、ブラシノステロイド受容体のBRI1と赤色蛍光タンパク質(RFP)との融合タンパク質形質転換体(BRI1-RFP)との二重形質転換体を作製したところ、BIL4-GFPとBRI1-RFPが重複して黄色シグナルとして観察されました(図6)。この結果と生化学的解析の結果を合わせた考察により、BIL4とBRI1は、エンドソームにおいて相互作用していることが分かりました。
さらに、BIL4はBRI1と相互作用することによって、どのような作用をBRI1にもたらしているか調べました。BRI1とGFPの融合タンパク質を持つ形質転換体(BRI1-GFP)と、BIL4が野生型よりも低発現している遺伝子組換え植物(BIL4低発現型形質転換体)の二重形質転換体を作製しました。すると、通常は細胞膜に多くみられるBRI1-GFPシグナルが、BIL4低発現型形質転換体では液胞内で多くみられました。このことから、BIL4の発現量が低下した条件下では、BRI1のエンドソームから液胞への運搬が進むことが分かりました。
以上の結果を整理したBIL4の機能モデルを図7に示します。左側のBIL4が発現していないときは、BRI1は細胞膜に主に局在していますが、一部はエンドサイトーシスで細胞内のエンドソームに取り込まれた後、液胞に運ばれて分解されます。BIL4は細胞伸長の極初期に限定的に発現することが、GUS融合タンパク質[10]を利用した解析によって明らかとなっています。右側のBIL4が発現している細胞では、BRI1のエンドソームから液胞への輸送と分解が妨げられます。その結果、細胞内のBRI1の量が増加し、ブラシノステロイドのシグナル伝達が活性化され、植物細胞の伸長が促されます。
すなわち、“細胞伸長の極初期に発現したBIL4が、BRI1の分解抑制を介して植物細胞の伸長を促すという仕組み”が存在することが明らかとなりました。
今後の期待
本研究によって得られたBIL4の遺伝子情報と知見を活用することによって、植物細胞の伸長を自在に制御する技術を開発することが可能になると考えられます。植物は、大気中の二酸化炭素を細胞壁など自らの体に固定するという生物活性を持っています。また、固定された炭素は植物バイオマスの原料となり、クリーンなバイオ燃料の源にもなります。さらに、途上国などで深刻化している食糧不足に向けて、穀物生産量の増大が求められています。
BIL4遺伝子は植物種を越えて、イネやトウモロコシ、サトウキビなどにも広く保存されている遺伝子です。これらの実用化作物におけるBIL4遺伝子の研究の発展は、地球環境の保護や食料増産などに貢献すると期待できます。
また、BIL4遺伝子は植物種のみでなく、ヒトやマウスなどの動物にも相同性の高い遺伝子が存在し、進化的に広く保存される遺伝子であることが分かっています。今後、動物におけるBIL4相同性遺伝子[11]の細胞伸長の機能についても解明が進むと期待できます。
原論文情報
- Ayumi Yamagami, Chieko Saito, Miki Nakazawa, Shozo Fujioka, Tomohiro Uemura, Minami Matsui, Masaaki Sakuta, Kazuo Shinozaki, Hiroyuki Osada, Akihiko Nakano, Tadao Asami and Takeshi Nakano, "Evolutionarily conserved BIL4 suppresses the degradation of brassinosteroid receptor BRI1 and regulates cell elongation.", Scientific Reports
発表者
理化学研究所
環境資源科学研究センター 機能開発研究グループ
専任研究員 中野 雄司(なかの たけし)
研究員 山上 あゆみ(やまがみ あゆみ)
グループディレクター 篠崎 一雄(しのざき かずお)
報道担当
理化学研究所 広報室 報道担当
Tel: 048-467-9272 / Fax: 048-462-4715
お問い合わせフォーム
産業利用に関するお問い合わせ
理化学研究所 産業連携本部 連携推進部
お問い合わせフォーム
補足説明
- 1.ステロイドホルモン
多細胞生物が種を越えて広く持っている生理活性化合物。ステロイド骨格と呼ばれる4員環骨格を持つ。哺乳類では筋肉などを作る男性ホルモンのテストステロン、妊娠の維持に関わる女性ホルモンのプロゲステロン、昆虫では脱皮に関係するエクダイソンが知られる。植物でもステロイドホルモンは存在し、ブラシノステロイドと呼ばれる化合物が植物の成長や葉緑体の発達を制御している。各生物種において細胞伸長や細胞分裂など、生物種を越えた共通の生理活性を持つと同時に、各生物種に固有の生理活性を持つ化合物。 - 2.ブラシノステロイド
アブラナの学名であるBrassica(ブラシカ)が名前の由来で、約10種類ある植物ホルモンの一つ。1979年にアメリカ農務省の研究グループがアブラナの花粉から発見し、化学構造を決定した。他の植物ホルモンが植物に固有の化合物であり、葉緑体内で生合成される酵素反応の段階があるなど植物に限定的であるのに対して、ブラシノステロイドは生合成経路の全てが細胞質で行われ、動物や昆虫にも類縁化合物が存在するなど生物種間で普遍的であるのが特徴。植物に対しては、細胞伸長や細胞分裂の促進など細胞レベルの促進的作用や、子葉の開化、胚軸の伸長、維管束の分化、緑葉の上偏成長、葉柄の伸長、茎の伸長など器官レベルの制御作用、また、ストレス耐性の付与(耐冷、耐塩、耐乾燥)や植物病害抵抗性の促進(植物自然免疫の活性化)など、さまざまな生理作用を示す。 - 3.ブラシナゾール(Brz)
ブラシノステロイド生合成経路を阻害する化合物。1998年に理研植物機能研究室の浅見忠男博士(現:東京大学大学院農学研究科)らが創製した。Brzを植物に処理すると、植物を生合成欠損変異体と同じような形・大きさにすることが可能である。植物の遺伝子を変異させてブラシノステロイド欠損状態にする従来の方法では、成長の最初から最後まで定常的に、ブラシノステロイド欠損状態を植物に与えることになる。しかし、Brzを使用することにより、全ての植物が、成長初期は通常量のブラシノステロイドがあるにも関わらず、その後のBrz処理によって、特定の成長時期以降に、特定の減少量でブラシノステロイド欠損程度を調整することができる。この手法により、植物成長におけるブラシノステロイドの生理作用をより詳細に明らかにすることが可能となった。 - 4.ケミカルバイオロジー
化学物質(ケミカル)の力により、生命の仕組み(バイオロジー)を明らかにする研究手法。生物の形態やタンパク質の働きに対して生理活性を示す小分子化合物を有機合成により人工的に作製し、それを用いて標的タンパク質分子の同定やタンパク質の機能解明を目指すことなどを行う。有機合成化学を出発点とする点で、タンパク質の機能解明により生物自身が体内で合成する天然物の働きを明らかにすることを主とする通常の生化学的手法とは区別して考えることが多い。 - 5.相同性タンパク質
タンパク質を構成するアミノ酸配列が共通の祖先を持つこと。 - 6.細胞内小器官
オルガネラともいう。細胞内の膜に囲まれた構造体で、それぞれが定められた機能を果たす。具体的には細胞核、小胞体、ゴルジ体、エンドソーム、リソソーム、ペルオキシソーム、ミトコンドリア、葉緑体、液胞など。 - 7.エンドソーム
細胞小器官の一つ。細胞のエンドサイトーシス[8]によって細胞内に取り込まれた、物質の輸送や代謝に関与する袋状の構造体。 - 8.エンドサイトーシス
細胞が細胞外の物質(低分子やタンパク質など)を細胞膜で取り囲み、細胞質内に取り込む作用のこと。細胞が取り込もうとする物質の多くは大きな極性分子であることから、疎水性の細胞膜を通過することができない。そのため細胞は、エンドサイトーシスによって物質を取り込む。 - 9.液胞
細胞内小器官の一つ。植物細胞ではその体積のほとんどを占めることがある。これまでは老廃物の貯蔵庫としての認識が主であったが、最近では独自の代謝系を持つことや、細胞全体の代謝に影響を与える働きなどが確認されつつある。 - 10.GUS融合タンパク質
植物で用いられるレポーター遺伝子の一つ。ベーターグルクロニダール(GUS)タンパク質との融合タンパク質を、研究対象とする遺伝子のプロモーター領域に接続したベクターを持つ形質転換植物と、染色試薬X-Gluを組み合わせた解析によって、遺伝子の発現器官が青色で染色されて観察できる。 - 11.相同性遺伝子
遺伝子を構成する塩基配列が共通の祖先を持つ遺伝子のこと。
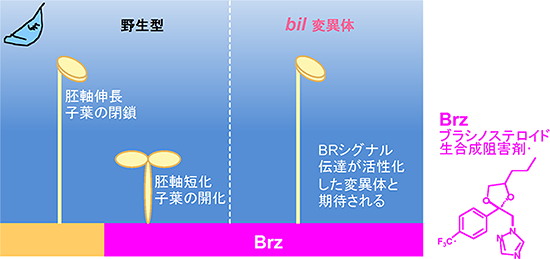
図1 bil4変異体のスクリーニング(探索)の原理
通常、野生型植物は暗所発芽条件で「もやし様」の徒長した形態を示す(左)。野生型植物を、暗所Brz培地で発芽させた場合、ブラシノステロイド欠損によって、暗所にも関わらず光条件下で発芽した植物のような胚軸が短化し、子葉が開化した形態の「暗所光形態形成」を示す(中央)。
暗所Brz条件下で胚軸徒長を示す変異体bil4(Brz-insensitive-long hypocoty-4l)が単離できれば、この変異体はブラシノステロイドのシグナル伝達が活性化したことによって、変異形態を獲得したと考え、スクリーニングを行った(右)。

図2 シロイヌナズナの野生型とBIL4が高発現した場合との草丈の違い
左)シロイヌナズナの野生型
中)BIL4が野生型よりも高発現している変異体
右)BIL4が野生型よりも高発現している遺伝子組み換え植物
BIL4が野生型よりも高発現しているシロイヌナズナは、野生型に比べて、草丈が長くなる傾向がみられた。

図3 シロイヌナズナの野生型とBIL4が低発現した場合の草丈の違い
左)シロイヌナズナの野生型
中右)BIL4が野生型よりも低発現している遺伝子組み換え植物
BIL4が野生型よりも低発現しているシロイヌナズナは、野生型に比べて、草丈が短くなる傾向がみられた。
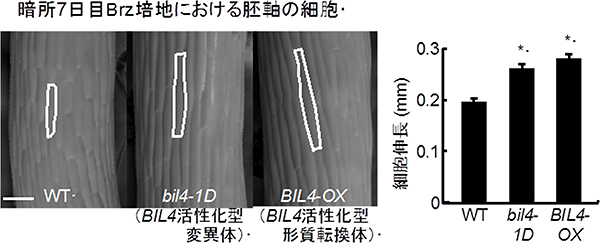
図4 シロイヌナズナの野生型とBIL4が高発現した場合の胚軸の表皮細胞の長さの違い
左)左からシロイヌナズナの野生型、BIL4が高発現している変異体、BIL4が高発現している遺伝子組換え植物の胚軸の表皮細胞の顕微鏡写真。
右)胚軸の表皮細胞の長さを示したグラフ。胚軸の表皮細胞の長さは、野生型に比べてBIL4が高発現した場合の方が長い。
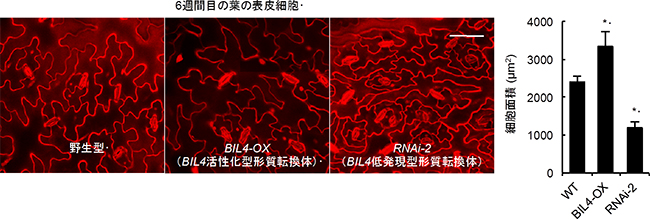
図5 シロイヌナズナの野生型とBIL4が高発現した場合と低発現した場合の葉の表皮細胞の違い
左)ロゼッタ葉の表皮細胞の顕微鏡写真。左からシロイヌナズナの野生型、BIL4が高発現している遺伝子組換え植物、BIL4が低発現している遺伝子組換え植物。
右)細胞面積を示したグラフ。細胞面積は野生型に比べて、BIL4が高発現している遺伝子組換え植物では大きく、BIL4が低発現している遺伝子組換え植物では小さい。
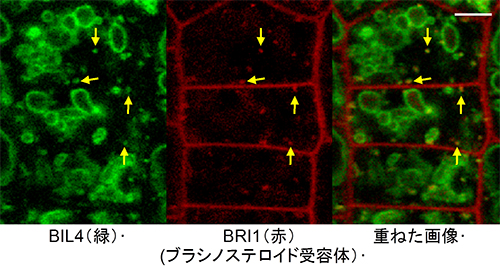
図6 BIL4とBRI1のエンドソームにおける相互作用
左)BIL4-GFPの画像。BIL4-GFPがドット状の明るい蛍光シグナルとして観察された。ドットはエンドソームであった。
中)BRI1-RFPの画像。
右)左と中を重ねた画像。BIL4-GFPとBRI1-RFPが重複して黄色シグナルが観察された。BIL4とBRI1は、エンドソームにおいて相互作用していることが分かった。
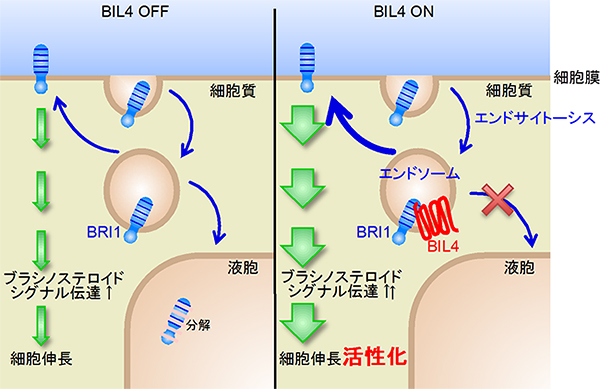
図7 BIL4によるBRI1の分解抑制モデル
左)BIL4が発現していないときは、BRI1は細胞膜に主に局在しているが、一部はエンドサイトーシスで細胞内のエンドソームに取り込まれた後、液胞に運ばれて分解される。
右)BIL4が発現している細胞伸長の極初期の細胞では、BRI1のエンドソームから液胞への輸送と分解が妨げられる。その結果、細胞内のBRI1の量は増加し、ブラシノステロイドのシグナル伝達が活性化され、植物細胞の伸長が促される。