2020年9月18日
理化学研究所
欧州分子生物学研究所
ポンペウ・ファブラ大学
京都大学
ヒトの時間ネズミの時間
-タンパク質の合成と分解速度が体節時計周期の多様性を生む-
理化学研究所(理研)生命機能科学研究センター再構成生物学研究ユニットの戎家美紀ユニットリーダー(研究当時、現欧州分子生物学研究所(EMBL) Barcelonaグループリーダー)、松田充弘研究員(研究当時、現EMBL Barcelona研究員)、ポンペウ・ファブラ大学のジョルディ・ガルシア教授、京都大学ウイルス・再生医科学研究所の影山龍一郎教授、京都大学iPS細胞研究所のジャンタシュ・アレヴ助教(研究当時、現京都大学高等研究院ヒト生物学高等研究拠点(ASHBi)准教授)、戸口田淳也教授、池谷真准教授らの国際共同研究グループは、ヒトの発生時間がマウスよりも遅いのは、遺伝子発現やタンパク質分解などの速度が、ヒトではマウスに比べて遅いことに起因することを発見しました。
本研究成果は、「ヒトとマウスの時間の違いがどのように生じるのか」という生物学上の根本的な問いを解き明かしたものです。
脊椎動物の発生期における重要イベントである「体節時計[1]」は、遺伝子発現の振動現象であり、規則的な体節形成の中心原理です。
今回、国際共同研究グループは、培養皿上で多能性幹細胞[2]から体節時計を再現し、マウスとヒトの体節時計周期の時間スケールが異なる理由を調べました。体節時計の中心となる遺伝子をヒトとマウスで入れ替えた細胞を作製したところ、マウスの遺伝子を持ったヒト細胞はヒトの時間(5時間周期)、ヒトの遺伝子を持ったマウス細胞はマウスの時間(2時間周期)を示しました。またこの違いは、タンパク質の分解速度や遺伝子発現の遅れなどの生化学反応が、ヒトの細胞ではマウスよりも2倍から3倍程度遅いために生じることが分かりました。すなわち、ヒトとマウスの時間の違いは体節時計遺伝子の違いではなく、細胞内環境の違いにより生じることが明らかになりました。
本研究は、科学雑誌『Science』(9月18日号)の掲載に先立ち、オンライン版(9月17日付:日本時間9月18日)に掲載されます。
背景
受精卵から体が作られる発生過程はヒトとマウスで非常によく似ており、器官ができる順番やその形成メカニズムはほぼ共通しています。しかし、妊娠期間はヒトが約9か月、マウスが約20日とヒトのほうが長く、寿命や心臓の拍動リズムもヒトのほうが長いことが知られています。これらの種に特異的な時間スケールは、どのように決まっているのでしょうか。国際共同研究グループは、発生の仕組みの一つである「体節時計」をモデルとして、ヒトとマウスの時間スケールの違いがどのように生じるかを明らかにしようと考えました。
体節は、背骨に代表される体幹部の繰り返し構造のもととなる組織で、体の中心軸の左右に規則的に形成される細胞の塊です。体節の数は発生に伴って増えていき、板状の細胞層(未分節中胚葉)が頭側から順番にくびれていくことで、左右1対の体節が体の後方に次々と付加されていきます(図1)。体節が1対増えるのにかかる時間は種によって異なり、ゼブラフィッシュ(魚類)は約30分、ニワトリ(鳥類)は約90分、マウスとヒト(哺乳類)ではそれぞれ約2時間と約5時間です。体節時計は、この体節形成に必須の役割を果たす原理で、その実態は遺伝子発現のオンとオフが周期的に切り替わる「振動現象」です。この時計の振動周期は体節形成の周期と一致し、マウスでは約2時間、ヒトでは約5時間であることが知られています(図2)。
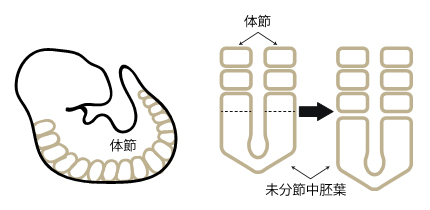
図1 脊椎動物胚の体節形成
- 左:マウス胚の体節の形状と位置を示した模式図。
- 右:体節の形成過程を背側から見た模式図。胚の後方に広がる板状の細胞層(未分節中胚葉)がくびれていくことで、新たな体節が付加される。
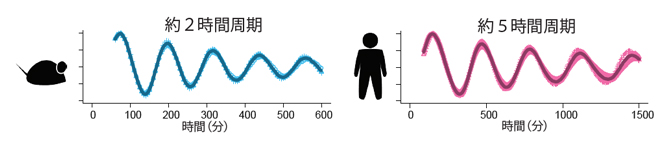
図2 マウスとヒトの体節時計
マウスでは約2時間、ヒトでは約5時間周期の遺伝子発現振動が観察される。 縦軸は、体節時計遺伝子発現の相対量。
体節時計の中心となるメカニズムは、転写因子[3]HES7をコードする「HES7遺伝子[4]」の自己抑制遺伝子回路であると考えられています。これは、HES7遺伝子の転写が、自身の遺伝子産物であるHES7タンパク質によって抑制されるという観察から導き出されたモデルです。すなわち、HES7遺伝子の転写が始まると、細胞内のHES7タンパク質が増加して自身の転写制御領域であるプロモーター[5]に結合し、その転写を抑制します。しばらくすると、HES7タンパク質が次第に減少し、HES7遺伝子の転写抑制が解除され、HES7タンパク質量が再び増加するという現象が繰り返されることで、時間的なリズムが生じると説明されます。マウスとヒトは、同じHES7遺伝子により体節時計を動かしていることが分かっています。では、どうして周期に違いがあるのでしょうか。
研究手法と成果
マウスとヒトの発生過程で見られる体節形成を、実際の胚で直接観察し、比較することは極めて困難です。国際共同研究グループは先行研究において、マウスとヒトの多能性幹細胞を用いて、体節時計を持つ組織を培養皿上で再構成する実験系を確立しました注1)。本研究ではこの系を用いて、マウスとヒトの体節時計を比較しました。
最初に、「マウスとヒトのHES7遺伝子領域の配列の違いが、周期の長さの違いを生む」という仮説を立てました。生物種間で異なるさまざまな特徴は、共通祖先種のゲノム配列に進化の過程で変異が入るために生じると考えられています。そこで体節時計において、中心遺伝子であるHES7遺伝子配列の種間差によって、マウスとヒトの時間の違いが生じたのかを検証しました。
実際に行った実験は、マウスとヒトのHES7遺伝子を交換するというものです。まず、ヒトHES7遺伝子を持ったマウス細胞と、マウスのHes7遺伝子を持ったヒト細胞を作製し、体節時計の時間を比べました。その結果、ヒトHES7遺伝子を持つマウス細胞は約2時間の周期を、マウスHes7遺伝子を持つヒト細胞は約5時間の周期を示しました(図3)。また、Hes7遺伝子をヒトHES7遺伝子に交換した遺伝子改変マウス胚の体節形成を観察したところ、その周期はやはり正常マウスに近い長さを示しました。これらのことから、体節時計の周期の違いはHES7遺伝子自体の違いではなく、反応の器である細胞の違いによることが分かりました。
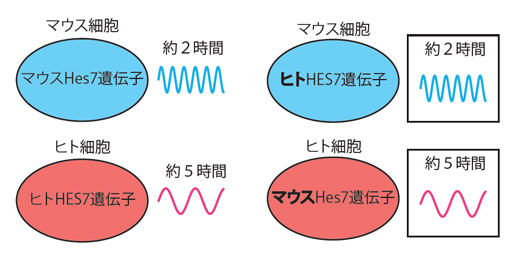
図3 ヒトの体節時計遺伝子を持ったマウス細胞とマウスの体節時計遺伝子を持ったヒト細胞
遺伝子改変により、ヒトの体節時計遺伝子(ヒトHES7遺伝子)を持ったマウス細胞とマウス体節時計遺伝子 (マウスHes7遺伝子)を持ったヒト細胞を作製し、体節時計の周期を測定した。その結果、前者は約2時間でマウスの時間、後者は約5時間でヒトの時間を示した。これらの結果から、体節時計の時間周期は体節時計遺伝子側ではなく、細胞側が決めていることが示された。
体節時計の中心原理は、HES7遺伝子の自己抑制遺伝子回路です。この遺伝子回路は、合成(発現)過程、抑制過程、分解過程という、三つの部分に分割することができます。どの過程がマウス細胞とヒト細胞で異なるのかを明らかにするために、それぞれの反応過程を測定し比較したところ、ヒトでは合成過程に要する時間が長く、HES7タンパク質の分解速度も遅いことが分かりました(図4)。これらの結果から、マウスとヒトでHES7遺伝子自体の機能は同じなのですが、細胞環境の違いによって、合成や分解といったHES7タンパク質の振る舞いに要する時間が違ってくることが示されました。
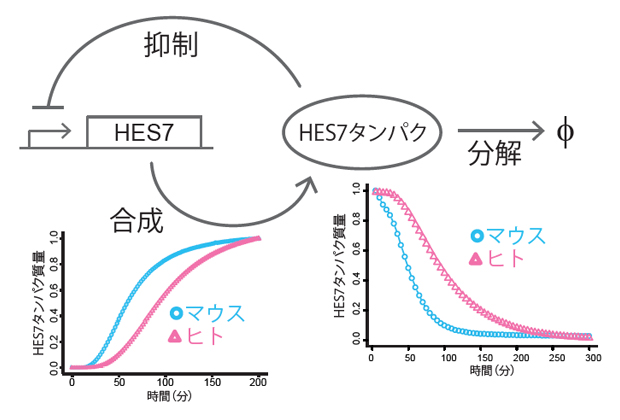
図4 体節時計の各プロセスの個別測定と種間比較
体節時計は合成(発現)と抑制と分解の三つのプロセスに分けられる。それぞれを測定し、マウスとヒトで比較したところ、合成過程と分解過程のいずれもヒトの方がマウスよりも遅いことが分かった。
最後に、今回の発見が体節時計に限定的なものであるのか、もしくは他の現象にも適用可能なのかを調べました。今回の発見は、「マウスとヒトでは、HES7遺伝子の発現に要する時間とHES7タンパク質の分解速度が異なることで、体節時計の時間スケールが異なってくる」というものです。そこで、発生過程に関わるさまざまな遺伝子の発現に要する時間とタンパク質分解速度を調べたところ、HES7以外の遺伝子でもマウスとヒト細胞で違いが見られました。また、体節時計を持つ細胞とは異なる種類の細胞である神経系の細胞を多能性幹細胞から誘導し、種に特異的な時間の違いが見られるのか調べたところ、そこでもやはりヒトでは合成に要する時間が長く、タンパク質分解が遅くなっていました。
以上の結果から、今回明らかになった時間スケールの種間差を生み出すメカニズムは、体節時計に限定されたものでなく、マウスとヒトで見られる生命現象の時間スケールの違いを広く説明する可能性があることが示されました。
- 注1)M. Matsuda, Y. Yamanaka et al., Recapitulating the human segmentation clock with pluripotent stem cells. Nature 580, 124-129 (2020).
今後の期待
本研究では、ヒトの時間がマウス時間よりも遅いのは、細胞環境の違いによって遺伝子発現に要する時間が長く、タンパク質分解が遅いためであることが分かりました。しかし、細胞環境のどの違いがこうした差を生み出すのかは全く分かっていません。
また、今回は時間に注目しましたが、ヒトとマウスを比べたときの著しい違いとして、体のサイズがあります。生物種を広く見渡したとき、時間と体のサイズには強い相関があることが知られており、今回の発見は、種間のサイズの違いがどのように生じるかという問いの理解にもつながると期待できます。時間や体のサイズといった多細胞生物の多様性を生み出す細胞環境の違いの解明が、次の課題となります。
補足説明
- 1.体節時計
体節になる前の未分節中胚葉の細胞が持つ、体節形成の周期性を支配する分子時計。遺伝子発現の振動を示し、その振動周期は生物種間で異なる。 - 2.多能性幹細胞
さまざまな種類の細胞に分化することができる細胞。本研究ではマウスのES細胞(胚性幹細胞)とヒトのiPS細胞(人工多能性幹細胞)を用いた。 - 3.転写因子
遺伝子の発現を調節するタンパク質。DNA上に存在する遺伝子のシス転写調節領域に結合し、DNAを鋳型としてRNAが産生(転写)される時期や量を調節する。 - 4.HES7遺伝子
転写を抑制する転写因子をコードする遺伝子。HES7遺伝子のプロモーターにはHES7タンパク質が結合する配列が存在し、自身の転写を抑制する。HESはhairy and enhancer of splitの略。 - 5.プロモーター
DNA上で遺伝子(RNA)として転写される領域の近くにあり、遺伝子を発現させる機能を持つ領域(配列)。HES7遺伝子のプロモーターにはN-box(CACNAG)と呼ばれる配列が存在し、HES7タンパク質はこの配列に結合することで、遺伝子発現を阻害する。
国際共同研究グループ
理化学研究所 生命機能科学研究センター
再構成生物学研究ユニット
ユニットリーダー(研究当時) 戎家 美紀(えびすや みき)
(現欧州分子生物学研究所(EMBL) Barcelona グループリーダー)
研究員(研究当時) 松田 充弘(まつだ みつひろ)
(現欧州分子生物学研究所(EMBL) Barcelona 研究員)
研究員(研究当時) 林 華子(はやし はなこ)
ポンペウ・ファブラ大学(Universitat Pompeu Fabra)
教授 ジョルディ・ガルシア(Jordi Garcia-Ojalvo)
京都大学
ウイルス・再生医科学研究所
教授 影山 龍一郎(かげやま りょういちろう)
教務補佐員(研究当時) 吉岡-小林 久美子(よしおか-こばやし くみこ)
(現高等研究院ヒト生物学高等研究拠点(ASHBi) 研究員)
iPS細胞研究所
助教(研究当時) ジャンタシュ・アレヴ(Cantas Alev)
(現高等研究院ヒト生物学高等研究拠点(ASHBi) 准教授)
博士課程学生(研究当時) 山中 良裕(やまなか よしひろ)
(現高等研究院ヒト生物学高等研究拠点(ASHBi) 研究員)
教授 戸口田 淳也(とぐちだ じゅんや)
准教授 池谷 真(いけや まこと)
研究支援
本研究は、武田科学振興財団、Spanish Ministry of Science, Innovation and Universities (MCIU) and co-funded by the European Regional Development Fund (ERDF, EU)、日本学術振興会(JSPS)科学研究費補助金基盤研究(C)「構成的アプローチによる体節時計の仕組みの解明(研究代表者:松田充弘)」、同新学術領域研究(研究領域提案型)「体節時計をモデルとしたマウスとヒトの時間スケール種差を生み出す原理の解明(研究代表者:松田充弘)」「振動遺伝子による時間制御機構(研究代表者:影山龍一郎)」、同挑戦的萌芽研究「ヒト多能性幹細胞からの腱・靭帯誘導法の開発(研究代表者:ジャンタシュ・アレヴ)」、内藤記念科学振興財団による支援を受けて行われました。
原論文情報
- Mitsuhiro Matsuda, Hanako Hayashi, Jordi Garcia-Ojalvo, Kumiko Yoshioka-Kobayashi, Ryoichiro Kageyama, Yoshihiro Yamanaka, Makoto Ikeya, Junya Toguchida, Cantas Alev, Miki Ebisuya, "Species-specific segmentation clock periods are due to differential biochemical reaction speeds", Science, 10.1126/science.aba7668
発表者
理化学研究所
生命機能科学研究センター 再構成生物学研究ユニット
ユニットリーダー(研究当時) 戎家 美紀(えびすや みき)
(現欧州分子生物学研究所(EMBL) Barcelonaグループリーダー)
研究員(研究当時) 松田 充弘(まつだ みつひろ)
(現欧州分子生物学研究所(EMBL) Barcelona研究員)
ポンペウ・ファブラ大学(Universitat Pompeu Fabra)
教授 ジョルディ・ガルシア(Jordi Garcia-Ojalvo)
京都大学
ウイルス・再生医科学研究所
教授 影山 龍一郎(かげやま りょういちろう)
iPS 細胞研究所
助教(研究当時) ジャンタシュ・アレヴ(Cantas Alev)
(現高等研究院ヒト生物学高等研究拠点(ASHBi)准教授)
教授 戸口田 淳也(とぐちだ じゅんや)
准教授 池谷 真(いけや まこと)
報道担当
理化学研究所 広報室 報道担当
お問い合わせフォーム
京都大学総務部広報課 国際広報室
Email: comms [at] mail2.adm.kyoto-u.ac.jp
※[at]は@に置き換えてください。