2009年2月24日
理化学研究所
酵母ミトコンドリアでは二重らせんをひねらずにDNA組み換えを開始
- 新原理の発見で、試験管内と生体内での仕組みの矛盾を解消 -
ポイント
- 酵母ミトコンドリアの酵素Mhr1が予測に反し、二重らせんをひねらずに相同対合
- 定説の相同対合とは異なる二重らせんの組み換えの反応機構と反応生成物を発見
- ミトコンドリア症や老化に深くかかわるヘテロプラスミーの抑制に新たな手がかり
要旨
独立行政法人理化学研究所(野依良治理事長)は、酵母ミトコンドリアのDNA組み換えに必要なMhr1タンパク質※1による相同対合※2が、予測に反して、二重らせんをひねらずに相同DNA組み換え(遺伝的組み換え)※3の中間体であるDNA鎖交換体を作るとともに、従来相同対合を促進するとされた二重鎖DNAの右巻きの超らせん※4が、組み換え反応を妨げることを発見しました。Mhr1による相同対合では、細胞核でのDNA組み換えに必須なRecA型タンパク質(RecA、Rad51、Dmc1)※5による相同対合とは異なる新規の反応機構が働き、その生成物もこれまで知られるDループ※6ではなく、三重鎖と考えられるDNA鎖交換体ができていることを発見しました。理研基幹研究所吉田化学遺伝学研究室の凌楓専任研究員と柴田遺伝制御科学研究室の柴田武彦上席研究員の研究成果です。
DNAは、10.5塩基対で1回の右巻きのねじれ(ツイスト)を持つ二重らせんです。複製、転写、相同DNA組み換えを行うためには、原理的に二重らせんのツイストを緩めなければならず、その結果DNA全体に、逆向きの左巻き方向のねじれ(超らせん)をため込みます。RecA型タンパク質は、DNAのツイストを緩めて、組み換え相手のDNA(単鎖)と二重鎖を作れる部分を探し出して、DループというDNA鎖交換体を作ることが、これまで30年来の内外の研究で定説となっています。試験管内では、この反応の仕組みにより、生体から取り出した天然の二重鎖DNAが持つ右巻きの超らせんが、相同対合で生じる左巻き超らせんを中和するため、RecA型タンパク質による相同対合を促進するとされていました。しかし一方、生体内では、クロマチン構造※7やDNAトポイソメラーゼ※8により超らせんに由来するDNA分子内のストレスが解消され、超らせんを持たないDNAとして振る舞うと考えられています。今回の発見は、ミトコンドリアに固有な相同対合タンパク質の特徴を明らかにしただけではなく、試験管内での相同対合の原理と生体内のDNAが持つ特性の間のギャップを埋める、相同DNA組み換えの仕組みを提示するものです。相同対合は、DNA組み換え、修復だけでなく、特にミトコンドリアでは、DNA複製に加えてミトコンドリア症※9や老化が深くかかわるヘテロプラスミー※10の抑制に、きわめて重要な働きをしていることが知られており、今回の成果はその仕組みの核心に迫るものといえます。
本研究成果は、米国の科学雑誌『Journal of Biological Chemistry』(5月号)およびJBC Onlineに掲載されます。
背景
DNAは、10.5塩基対で1回のねじれ(ツイスト)を持つ右巻きの二重らせん構造をしています。一方、DNA自身の複製、DNAを鋳型とする転写、別のDNAとの間でDNA鎖を交換する相同DNA組み換えを行うためには、原理的に二重鎖DNAのツイストを緩める必要があります。ツイストが緩むと、必然的にDNA全体に、左巻き方向のねじれ(超らせん)をため込むことになります。
相同DNA組み換えの開始段階では、別の二重鎖由来の単鎖DNA末端が二重鎖DNAに割り込んで、その相補鎖と対合したDNA鎖交換体という組み換えの中間体を作る「相同対合」という反応が起こります(図1)。研究チームは、酵母ミトコンドリアで相同対合を行う酵素「Mhr1」が、ミトコンドリアDNA(mtDNA)の相同DNA組換えと複製に加えて、ミトコンドリア症に深くかかわるヘテロプラスミーの抑制やホモプラスミー※10化で、鍵となる働きをすることを明らかにしてきました(RIKEN NEWS No. 275 2004年5月号 研究最前線、2006年12月5日プレス発表)。
一方、細胞核では、相同対合を行うRecA型タンパク質(RecA、Rad51、Dmc1)が、発がんの抑制や正常な配偶子(卵子や精子)の形成で、きわめて重要な働きをしています。RecA型タンパク質による相同対合は、アデノシン三リン酸(ATP)※11を必要としますが、Mhr1や細胞核のRad52タンパク質による相同対合にはATPは不要です。そこで、RecA型タンパク質による相同対合反応と、Mhr1やRad52による相同対合反応の共通点と違いが専門家の間で議論されていました。
RecA型タンパク質は、親となる二重鎖DNAのツイストを緩めて、組み換え相手の単鎖DNAとの相補塩基配列部分を探し出し、分子間でDNA鎖を交換してDNA鎖交換体を作ります。親の二重鎖DNAの一方の鎖は、交換の結果相手を失うため、親の2つのDNA鎖の間のツイストは緩んだままとなり、その形からDNA鎖交換体は「Dループ」と呼ばれます。この反応の仕組みは、1979年にRecAが相同対合活性を持つことが発見されて以来の研究で定説となっています。
右巻きの超らせんを持つ天然の二重鎖DNAは、二重鎖DNAのツイストを緩めると生じる左巻きの超らせんを中和します。また、できたDループが、右巻きの超らせんを解消した状態を保つことになります(図2)。事実、試験管内では天然の二重鎖DNAの右巻き超らせんは、RecA型組み換え酵素によるDループ形成を大いに促進します。しかし、生体内、特に真核生物の核では、天然の二重鎖DNAが持っている右巻きの超らせんによるDNA分子内のストレスが、クロマチン構造を作ることで解消されます。また、核とミトコンドリアには、超らせんによるDNA分子内のストレスを解消する酵素のDNAトポイソメラーゼが存在します。従って、生体内のDNAは、核とミトコンドリアのいずれでも、超らせんを持たないDNAとして挙動すると考えられています。このように、試験管の中で行う組み換え反応での右巻き超らせんの働きと、生体内のDNAらせんが持っている特性との間には、大きなギャップがありました。
研究手法と成果
研究チームは、酵母ミトコンドリアの組み換え酵素「Mhr1」を単離し、試験管内で相同対合を行いました。その結果、理論的に反応を促進すると考えられていた右巻き超らせんが、相同対合反応を妨害していることが分かりました。また、反応が進むと左巻きの超らせんをため込むことで反応を妨害すると考えられていた超らせんを持たない閉環状二重鎖DNA(2つのDNA鎖がともに環になっている)でも、超らせんをため込まない環境での反応と同じ程度に相同対合が起こることを見つけました(図3)。
次に、Mhr1による相同対合反応の中間体と反応後のDNAを、DNAトポイソメラーゼで処理してゲル電気泳動を使い、DNAの立体位相幾何学的な状態を解析しました。この解析では、感度の高い分解能が得られるため、DNAの2つのねじれ(ツイストと超らせん)の状態を特定することができます※12。Mhr1による相同対合反応でできるDNA鎖交換体では、親の二重鎖DNAの2本のDNA鎖間のツイストが、外見上まったく緩んでいないという、これまで知られていなかった構造を示すことを発見しました(図4)。
この不思議な挙動の研究を進めた結果、Rad52へ結合した二重鎖DNAの構造的特徴(J. Biol. Chem., 283, 24264-24273 (2008))にヒントを得て、RecA型タンパク質による相同対合を促進する右巻き超らせんが、Mhr1による相同対合には必要ないばかりか、かえって反応を妨害するという相反する特性を矛盾なく説明できる、新たな相同対合の仕組みを明らかにしました(図5)。この仕組みでは、Rad52に結合した二重鎖DNAと同様に、二重鎖DNAがMhr1タンパク質の周囲へ右巻きに巻きつくことによって、二重らせんのツイストが緩んだ際にできる左巻きの超らせんが解消されます。その結果、外見上、二重鎖DNAのツイストも超らせんも変化していないように見えました。この仕組みでできるDNA鎖交換体は、対合した単鎖DNAと二重鎖DNAとの間でワトソン・クリック型塩基対ができ、単鎖DNAに置き換えられたDNA鎖が水素結合で結合した並行三重鎖構造に合致します。
今回発見したATPを必要としないMhr1による相同対合の仕組みは、ミトコンドリアの中のDNAでの相同対合に都合がよいだけではなく、RecA型タンパク質が試験管の中で行う組み換え反応での右巻き超らせんの働きと、生体内のDNAらせんが持っている特性とが一致しない理由を提示するものといえます。
今後の期待
研究チームは、これまでに、酵母ミトコンドリアにはRecA型タンパク質が存在せず、相同DNA組み換えの相同対合ではミトコンドリアタンパク質のMhr1が働くことを明らかにしていました。さらに、Mhr1タンパク質による相同対合が、ヘテロプラスミーからホモプラスミーへの復帰で重要な働きをし、その発現を制御することでホモプラスミー復帰の速さを制御できることも明らかにしています。今回、このMhr1タンパク質が、既知のRecA型タンパク質による相同対合と異なる機構で相同対合を行うことを発見し、ヒトのミトコンドリアにもあると予測されている相同対合タンパク質を同定する大きな手掛かりを得ました。
ヒトでは、ミトコンドリアDNAの相同的組み換えはないといわれていましたが、近年、ヒトでもミトコンドリアDNAの相同組み換えが起きていることを示す結果が集まってきています。一方、ヘテロプラスミーが深くかかわっているミトコンドリア症は、糖尿病などとの関連も認識され始めてきた重要な疾患ですが、その治療の手掛かりは極めて乏しい状況です。ミトコンドリア症の抑制には、ホモプラスミー状態への復帰が鍵と考えられていますが、ヒトではその仕組みも、関与する遺伝子やタンパク質もまったく分かっていません。ヒトのミトコンドリアにも、酵母ミトコンドリアと同様にRecA型タンパク質が存在しません。今回研究チームが明らかにした、酵母ミトコンドリアのMhr1による相同対合活性の特徴を手がかりに、ヒトミトコンドリアの相同対合を行うタンパク質を突き止めることができると、酵母と同様なホモプラスミー復帰の機構がヒトでも働いていることが明らかとなり、ヘテロプラスミー化を抑制し、ホモプラスミー化を促進することによって、ミトコンドリア症の治療につながる可能性を開くことになります。
発表者
理化学研究所
基幹研究所 柴田遺伝制御科学研究室
上席研究員 柴田 武彦 (しばた たけひこ)
Tel : 048-467-9528 / Fax : 048-462-1227
吉田化学遺伝学研究室
専任研究員 凌 楓 (りん ふぉん)
Tel : 048-467-9518 / Fax : 048-462-4676
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.Mhr1タンパク質
初めて単離されたミトコンドリアDNA(mtDNA)の組み換え欠損変異の責任遺伝子として同定された酵母の核染色体の遺伝子 MHR1( Mitochondrial Homologous Recombina-tion-deficiency, the 1st mutant)がコードするタンパク質。ATPを必要とせずに相同対合を行う活性を持つ。アミノ酸配列の類似性を基にしたヒトのMhr1探索は成功していないため、機能の同一性を基に探すための新たな手掛かりが必要であった。 - 2.相同対合
二重鎖切断の末端に由来する単鎖DNAが、同じ塩基配列を持つ別の二重鎖DNAの二重らせんに割り込んで、その相補鎖と対合する相同DNA組換えに特有な反応。 - 3.相同DNA組み換え
生物界で普遍的に観察される、一組の塩基配列が同じか、よく似ているDNA分子の間で遺伝情報を再編成する遺伝現象。RNAウイルスでは、相同DNA組み換えはむしろ稀。動物のミトコンドリアDNAについては、相同DNA組み換えは起こらないといわれていたが、最近、組み換えが起こったことを示す結果が集まってきた。相同DNA組み換えには、一方のDNAの塩基配列で相手の塩基配列が置き換えられるジーンコンバージョンと、互いに塩基配列を交換する交差(交叉)とがある。生体では、ジーンコンバージョンによるDNA二重鎖切断の修復と、配偶子(精子、卵子、胞子)を作る倍数体から半数体ができる細胞分裂(減数分裂)で、一対ずつある相同染色体の正確な分離と両親の遺伝形質を再編成することによる遺伝的な多様性の獲得が、その機能として知られている。 - 4.超らせん
2本のDNA鎖がともに環になっている閉環状二重鎖DNAでは、右巻きの二重らせん(ツイスト)に加えて、DNA全体のねじれが起こる。このDNA全体のねじれを超らせんという。これは、二重らせんがもつ固有のツイスト数と、2本のDNA鎖が互いにねじり合う回数(リンキング数)との間にずれがあるため、その差が超らせんとなって現れる。生体からの単離された閉環状二重鎖DNAは、そのリンキング数が二重らせんのツイスト数より小さいため、右巻きの超らせんを持っており、その二重らせんには、ツイストを減らす方向にストレスがかかっている。右巻き超らせんによる二重らせんにかかるストレスは、DNAを軸の周りに左巻きに巻きつけることで解消する。細胞核内のDNAは、ヒストンコアの周りに左巻きに巻きついているため、超らせんに由来するストレスはかかっていない。 - 5.RecA型タンパク質(RecA、Rad51、Dmc1)
1979年に研究チームは、大腸菌のRecAタンパク質が、ATP依存して相同対合を行い、Dループを作る活性を持つことを発見した。その後、広く真核生物が核に持つRad51タンパク質やDmc1タンパク質が、RecAと共通の構造を持ち、ATPを必要とする相同対合活性があることが1992年から1994年にかけて発見された。RecAタンパク質は、相同対合ではATPの加水分解は必要としないが、相同対合の後、ATPを加水分解して得られるエネルギーを用いて、ヘテロ二重鎖を特定方向へ伸長させる。 - 6.Dループ
二重鎖DNAで相同対合が起こった結果、外から取り込まれたDNAによって置き換えられたDNA鎖が対合の相手を失ってできるループ構造。Mhr1でも、右巻きの超らせんを持つ二重鎖DNAをつかって相同対合を行うと効率は下がるが、Dループを作る。この場合のDループは、おそらく、三重鎖と考えられる相同対合の生成物が、Mhr1タンパク質を除いた後に、DNAの持つ超らせんのストレスによって、非酵素反応として三重鎖の第3のDNA鎖が二重鎖から分離して生じる - 7.クロマチン構造
真核生物の核染色体DNA(約150塩基対)が、ヒストンコアの周囲に左巻きに1と3分の2回転巻き付いた構造。電子顕微鏡で観察すると数珠のように見える。ヒトでは全長が2mにもなるDNAを、10ミクロン程度の直径しかない細胞核内にコンパクトに収めるための最下層の構造。 - 8.DNAトポイソメラーゼ
二重鎖DNAの一方の鎖(I型)または、両方の鎖(II型)を一過的に切断して、DNAの超らせん状態を変える酵素。ATP存在下で、閉環状二重鎖DNAに右巻き超らせんを導入するDNAジャイレース以外は、閉環状二重鎖DNAが持つ超らせんを解消する。 - 9.ミトコンドリア症
ミトコンドリアDNAにコードされている固有のtRNAやポリペプチドの変異、または核染色体の変異により引き起こされる、酸化的リン酸化に働くタンパク質複合体の機能異常で起こる病気の総称。最近、糖尿病などとの関係が明らかになり、従来考えられていた以上の広がりを持つ疾患であることが分かってきた。 - 10.ヘテロプラスミー、ホモプラスミー
酵母や哺乳類の細胞は、一つの細胞に100弱から数万コピーのミトコンドリアDNA(mtDNA)を持っている。増殖している酵母や生まれたての健常人では、すべての細胞のmtDNAは同じ塩基配列(遺伝子型)を持つ。この状態をホモプラスミーと呼ぶ。変異体など異なる塩基配列をもつmtDNAが、一つの細胞または個体の中で共存している状態は、ヘテロプラスミーという。ミトコンドリア症の患者ではヘテロプラスミーが広く認められる。また、健常人でも、神経や筋肉では加齢とともにヘテロプラスミーが顕著になっていく。 - 11.アデノシン三リン酸(ATP)
生体でエネルギーを必要とする反応に、三リン酸が二リン酸、または一リン酸への加水分解と共役することで、エネルギーを供給する。 - 12.DNAのねじれの電気泳動による解析
タンパク質との結合やDループの形成で、二重らせんのねじれ(ツイスト)が緩んだ試料の閉環状二重鎖DNAをDNAトポイソメラーゼで処理して、その状態で持つ超らせんを解消する。その後、DNAトポイソメラーゼの活性を抑えてから、タンパク質や単鎖DNAを試料の閉環状二重鎖DNAから外すと、ツイストは二重鎖DNA固有の状態に(10.5塩基対で右巻きに1回)に戻るため、タンパク質との結合などで緩んでいた数と同数の右巻きのツイストが増えることになる。その結果、増えた右巻きツイストと同じ数だけの右巻き超らせんが導入される。その試料を、2次元の電気泳動で、対照となる超らせんを持たない閉環状二重鎖DNAと並べて泳動する。試料の閉環状二重鎖DNAの帯のセットと、対照の閉環状二重鎖DNAとの間にあるはずの帯の数を数えることで、試料の閉環状二重鎖DNAが持つ超らせんの数が正確に分かる。
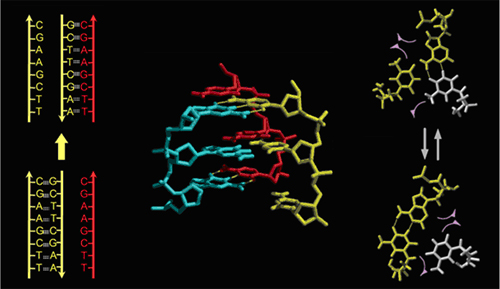
図1 相同対合による組み換えの中間体(DNA鎖交換体)の形成
(左) 矢印の方向に反応が進むにつれて親の二重鎖DNAの塩基対(黄)が、組み換え相手の単鎖DNA(赤)との間の塩基対にスイッチしてDNA鎖交換体ができる。
(中央と右) DNAは右巻きらせん構造をとっているために、塩基が水平回転することで相同対合を行う。この塩基の回転の自由度を確保するために、塩基対の間隔を広げる必要がある。塩基対の間隔を広げることと、二重らせんのツイストを緩ませ、全体を左巻きにねじりあげて、左巻きの超らせんを入れることとが連動する。(図の一部Nishinaka, T., Shinohara, A., Ito, Y., Yokoyama, S. and Shibata, T., Proc. Natl. Acad. Sci. USA, 95, 11071-11076 (1998) から引用)
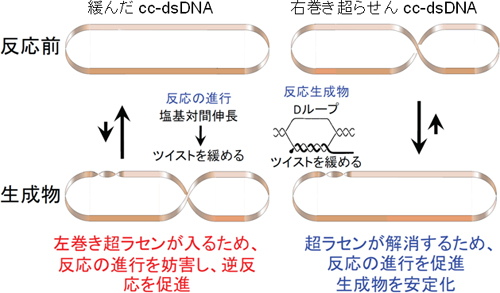
図2 これまで知られていた二重鎖DNAの超らせんと相同対合の関係
環状の帯は右巻き二重らせんを示す。ツイスト(DNAの右巻きらせん)の緩みは、帯を左巻きにひねることに対応する。相同対合の進行に伴いツイストが緩むと、もともと超らせんを持たない緩んだ閉環状DNA(2本のDNA鎖がともに環になっている、cc-dsDNA)では(左上)、左巻きの超らせんが入る(左下)ため反応の進行が妨げられる。また、反応でできるDループは、左巻き超らせんが解消するストレスがかかっていて不安定であり、超らせんの解消にともない、壊れてしまう。一方、もともと右巻きの超らせんを持つ天然のcc-dsDNA(右上)では、反応の進行とともに右巻き超らせんが解消する(右下)ので、反応の進行を促進し、できるDループは安定化する。RecA型タンパク質による相同対合は、この原理から期待される通りに起こる。
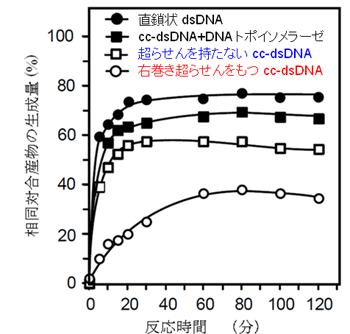
図3 右巻き超らせんはMhr1による相同DNA対合を妨害する
ツイストの変化が自由にできる直鎖状二重鎖DNA(dsDNA)やツイストの変化で生じる超らせんを解消するDNAトポイソメラーゼ存在下で、最も効率よくMhr1による相同対合が進行する。図2で示したようなツイストの変化で直ちに超らせんを生じてしまい、相同対合の進行を妨害すると考えられていた超らせんを持たない閉環状二重鎖DNA(cc-dsDNA)でも、上記の2種のDNAとほとんど同じ程度にMhr1による相同対合が起こる。一方、RecA型タンパク質による相同対合を促進する右巻き超らせんを持つ閉環状二重鎖DNAでは、Mhr1による相同対合が著しく妨害される。このように、Mhr1による相同対合はこれまでの知識で予測されていた結果(図2)とはまったく逆な結果になることを発見した。
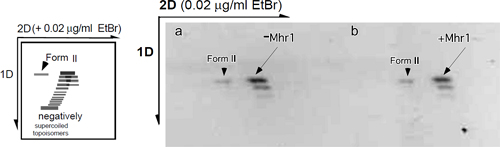
図4 Mhr1による相同対合でできた、ツイストの緩みがない反応生成物
DNAトポイソメラーゼ存在下で行ったMhr1による相同対合の生成物から、タンパク質と対合した単鎖DNAを除いた後、電気泳動でその超らせんの状態を解析することで、生成物中のツイストの状態を解析した。Mhr1による相同対合の生成物(+Mhr1)は、超らせんを持たない閉環状二重鎖DNA(-Mhr1)と同じ位置に二重鎖DNAの信号(帯のセット)が検出できた。この結果は、もとの二重鎖DNAのツイストをまったく変えない生成物ができたことを示し、Mhr1による相同対合ではDループを作らないことが分かった。最も可能性が高い生成物は、対合した単鎖DNAと二重鎖DNAとの間でワトソン・クリック型塩基対ができ、その主溝に置き換えられたDNA鎖が水素結合で結合した並行三重鎖である。電気泳動マーカーのFormIIは、DNA鎖に切れ目が入って超らせんが失われた環状二重鎖DNAで、閉環状二重鎖DNAの試料に必ず含まれるので、電気泳動のマーカーとなる。
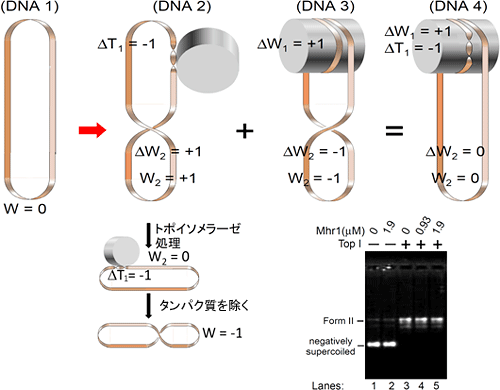
図5 超らせんを持たない閉環状二重鎖DNAの上で、外見上超らせんとツイストが変化しないで起こる相同対合のモデルとその仮説を支持する実験結果
RecA型タンパク質では、相同対合の途中で二重らせんのツイスト(T)を緩める(ΔT < 0)。ツイストを緩めると、外に左巻き超らせん(W > 0)を誘導する(DNA2)。Mhr1と同様にATPを必要とせずに相同対合を行うRad52では、そのタンパク質が作るリングに沿って二重鎖DNAが右巻きに巻きつく。タンパク質の周りに右巻きに巻きつくと、外には右巻きの超らせんを生じる(DNA3)。そこで、Mhr1では、DNAがタンパク質の周りに右巻きに巻きつくに従ってツイストが緩むと仮定すると、ツイストを緩めることで外に左巻き超らせんが生じて、超らせん同士が解消し合い、見かけ上二重鎖DNAのツイストと超らせん状態が変わらないことになる(DNA4)。DNA2の状態でDNAトポイソメラーゼ処理を行うと、ツイストが減少した状態で超らせんが解消される(W = 0)。その後タンパク質を除くと二重らせんのツイストが元に戻り(ΔT = 0)、今度はDNA全体に右巻きの超らせんが導入される(W < 0;下向きの矢印の過程)。一方、DNA4ではDNAトポイソメラーゼ処理をしても外見上何の変化も起こらない(写真のLane4、lane 5と対照のlane 3 (Mhr1なし)との比較)。
(参考)DNAの右巻き二重ラセンのねじれ(ツイスト)の数をT、超らせん(軸の周りへの逆向きの巻きつきと等価)の数をWで示す。天然の超らせんは右巻きで、負の値で示す(W < 0)。ツイストTの値は測定できないので、二重鎖DNAが標準状態で示すツイストとの差(ΔT)でツイストの変化を表す。ΔT + Wはトポイソマー(互いに超らせんの状態だけが異なる閉環状二重鎖DNA)それぞれに固有の整数値をとる。DNAトポイソメラーゼが働くと、この整数値が、W = 0 となるように変化する。ここでは、ΔT = -1(二重らせんの右巻きツイストを1回転だけ緩める、言い換えると二重鎖DNAを1回転左にひねる)の場合を代表して示している。実際に、紙テープで図のような輪を作って操作すると、ツイスト、超らせん、軸の周りの巻き付きの関係が容易に理解できる。