2009年3月10日
理化学研究所
アブラムシは、かつて別の細菌から獲得した遺伝子で必須共生細菌を制御する
-世界初、動物-細菌間の共生維持に利用される遺伝子の驚くべき起源を発見-
ポイント
- 細菌と動物の間の垣根を越えた、遺伝子資源の共有を確認
- 共生細菌のオルガネラ化など、共生に基づく生物進化に重要な示唆
- 昆虫の必須共生系の基盤理解で、安全な害虫防除法の開発に期待
要旨
独立行政法人理化学研究所(野依良治理事長)は、農業害虫として知られるアブラムシが、かつて感染していた細菌などから複数の遺伝子を獲得し、これらを発現させて、自身の生存に必須である相利共生細菌※1を維持・制御している、という証拠を、世界で初めて突き止めました。これは理研基幹研究所(玉尾皓平所長)宮城島独立主幹研究ユニットの中鉢淳ユニット研究員と放送大学(石弘光学長)二河成男准教授による共同研究の成果です。
アブラムシは、栄養分に乏しい植物の師管液だけを餌としながら、旺盛な繁殖力を示しますが、これを可能にしているのが、細菌との共生関係です。アブラムシは体腔※2内に、「菌細胞※3」と呼ばれる特殊な細胞をもち、この細胞質の中に、師管液に欠けている栄養分を合成する共生細菌「ブフネラ※4」を収納しています。アブラムシは、ブフネラから供給されるこの栄養分に依存して成育しており、ブフネラなしでは繁殖できません。一方、ブフネラは、ミトコンドリアや葉緑体などのオルガネラ(細胞小器官)と同様、アブラムシの親から子へと、1億年以上にわたって垂直感染※5だけで伝えられており、この過程で多くの遺伝子を失っているため、菌細胞の外では増殖できなくなっています。このように、アブラムシとブフネラは、もはや不可分で、ひとつの生物として振る舞う複合体を形成しているといえます。
研究グループは、アブラムシの菌細胞で特異的に高発現している遺伝子のうち、少なくとも2種類が、細菌からの水平転移※6によってアブラムシのゲノムに組み込まれたものであることを明らかにしました。その内の1つは、細菌の細胞壁のリサイクルにかかわるもので、驚くべきことにブフネラ由来ではなく、かつてアブラムシに感染していたと思われる別の細菌に由来することが分かりました。ブフネラは、進化の過程でこれら2種類の遺伝子のいずれも失っているため、こうした遺伝子が、アブラムシの菌細胞だけで高発現している事実は、2つの遺伝子がブフネラの遺伝子の欠如を補い、ブフネラの生存を支えていることを強く示唆します。
この成果は、細菌から動物ゲノムに水平転移したDNA断片が、遺伝子として機能できることを明確に例証するだけでなく、さらに水平転移した遺伝子が、転移元以外の細菌との共生関係に利用されうることを世界で初めて示すものです。これは、真核生物※7の細胞内にすむ細菌の進化動態に関する予想を覆すもので、そのオルガネラ化※8メカニズムの解明に貢献するばかりでなく、生物間の相互作用を扱う、基礎から応用にわたる幅広い分野に、大きなインパクトを与えるものです。
本研究成果は、英国のオンライン科学雑誌『BMC Biology』(3月10日付:日本時間3月10日)に発表されます。
背景
農業害虫として知られるアブラムシ(図1)は、必須アミノ酸などの栄養分が乏しい植物の師管液だけを餌としながら、きわめて旺盛な繁殖力を示します。これを可能にしているのが、微生物との緊密な共生関係です。アブラムシは、体腔内に「菌細胞」とよばれる特殊な細胞を持ち、この細胞質中に、大腸菌などに近縁な共生細菌で、必須アミノ酸やビタミンなどを合成する「ブフネラ」を多数収納しています。アブラムシは、ブフネラから供給される、こうした栄養分に成育を依存しているため、ブフネラの存在がないと繁殖することができません。
このブフネラは、元来、アブラムシの消化管内にすんでいた細菌に由来する、と考えられていますが、1億年以上前に体腔内に侵入した後は、アブラムシの親から子へと垂直感染のみによって受け継がれています。この特異な進化の過程で、ブフネラは、細菌の生存に必要と思われる多くの遺伝子を失っており、もはや菌細胞の外では増殖することができなくなっています。つまり、宿主アブラムシ、ブフネラともに単独では生存できず、ひとつの生物として振る舞う複合体を形成しているといえます。
それでは、宿主であるアブラムシの菌細胞は、どのような遺伝子を発現して、遺伝子数の乏しいブフネラの生存を支えているのでしょうか?これを調べるために中鉢博士らは、先行研究において、菌細胞で発現している遺伝子の網羅的解析を行いました (Nakabachi et al., PNAS, 2005)。その結果、アブラムシのゲノムにコードされていながら、細菌の遺伝子とよく似た遺伝子を2種類見つけました。ミトコンドリアや葉緑体といったオルガネラは、その進化過程で、自分の遺伝子を宿主ゲノムへ転移させてきました。この2種類の遺伝子も同様に、ブフネラからアブラムシへ水平転移したものなのでしょうか?今回の研究では、さまざまな手法を用いてこれらの遺伝子について解析し、その構造や機能、進化の歴史を明らかにしました。
研究手法と成果
はじめに、先行研究に用いたcDNAライブラリーから、2種類の遺伝子に対応するクローンをすべて拾い出し、これらの全長の塩基配列を決定しました。この情報に基づいて、あらためて転写産物や、アブラムシゲノム上の対応する遺伝子をクローニングし、塩基配列やエクソン/イントロン※9構造を確認しました。さらに、得られた塩基配列をタンパク質のアミノ酸配列に翻訳し、データベース上の類似タンパク質の情報と比較してドメイン構造を抽出した結果、タンパク質の機能を推定することができました。同時に、翻訳したアミノ酸配列を用いて分子系統解析※10を行い、遺伝子の進化的由来を推定しました。
また、遺伝子の発現レベル、発現部位の特異性について調べるため、リアルタイム定量RT-PCR法※11で、転写産物の存在量を菌細胞とアブラムシの体全体で測定し、両者の間で比較しました。
その結果、解析に用いた2種類の遺伝子のうちの1つは、細菌の細胞壁を構成するムレイン※12を代謝、リサイクルするために必要な酵素の「LD-カルボキシペプチダーゼ」をコードする「ldcA遺伝子」であると分かりました(図2A)。また、この遺伝子が、菌細胞で特異的に強く発現していることも分かりました(図3)。菌細胞に収納されているブフネラは、ムレインを持つものの、その代謝に必要なldcA遺伝子を欠いています。こうした遺伝子の発現が菌細胞で亢進している事実は、アブラムシのldcA遺伝子が、ブフネラのldcA遺伝子の欠如を補い、ブフネラの生存を支えている可能性を強く示唆するものです。
さらに驚くべきことに、ldcA遺伝子はブフネラの祖先種から水平転移したものではなく、「ボルバキア※13」と呼ばれる、ブフネラとは遠縁の「リケッチア※14」の仲間に近縁な細菌に由来するものであることが、分子系統解析から分かりました。ボルバキアやリケッチアは、昆虫を含む多くの節足動物から検出されていますが、今回解析に用いた系統のアブラムシには感染していないことを確認しています。つまりアブラムシは、かつて感染していたボルバキア様細菌から、水平転移によりこの遺伝子を獲得し、その細菌がアブラムシから失われてしまった後も、ブフネラの維持、制御に利用していると考えることができます。なお、ボルバキアに由来すると考えられるDNA断片は、これまでも複数の昆虫ゲノムから見つかっていましたが、これらが「遺伝子」として働いている証拠は、今回世界で初めて得られました。
もう1つの遺伝子は、「レアリポプロテインA」と呼ばれるタンパク質をコードする「rlpA遺伝子」でした(図2B)。このタンパク質の機能は未だ不明ですが、何らかの酵素機能を持つことが示唆されています。このrlpA遺伝子についても、ブフネラのゲノムには存在せず、さらに菌細胞で特異的に発現が亢進している(図3)ことから、やはりブフネラの生存に重要な役割を果たしていると推察されます。今回の解析では、この遺伝子を供給した細菌種を特定することはできなかったものの、アブラムシのレアリポプロテインAは、細菌由来の原核生物型主要ドメイン構造に加えて、真核生物型のシグナルペプチドや複数のモチーフ構造などからなるキメラ状の構造を持つことが分かりました。これは、細菌から水平転移した原核生物型の配列と、真核生物型であるアブラムシ本来の配列が融合することで形成されたものと推定されます。
なお、ldcA、rlpAのいずれの遺伝子にも、真核生物の遺伝子に特徴的で、細菌の遺伝子には見られない構造である、スプライソソーム型イントロン※9が、存在していました(図2)。また、これらの転写産物は、やはり真核生物に特徴的な5´-キャップ構造や3´- ポリA構造を持つことが示唆されています。これらのことから、原核生物である細菌の遺伝子が、真核生物であるアブラムシのゲノムに取り込まれた後、真核生物型の遺伝子構造、転写産物構造を獲得したことが分かります。
本研究から、アブラムシは、細菌から複数の遺伝子を水平転移により獲得し、これを自らの生存に必須である相利共生細菌の維持、制御に利用しているという証拠を得ることができました。これまでも、細菌に由来すると思われるDNA断片が動物のゲノムから見つかることはありましたが、原核生物と真核生物では遺伝子の発現機構が異なるため、こうしたDNA断片のほとんどは「遺伝子」として機能することなく、壊れていく一方だと考えられていました。今回の研究は、細菌ゲノムから動物ゲノムに水平転移したDNA配列が、実際に遺伝子として機能できることを明確に例証したばかりでなく、こうした遺伝子が、さらにほかの細菌との共生関係に利用されているという前代未聞の新知見をもたらしました。この事実は、生物がきわめて柔軟性に富むことを示しており、生物間の相互作用を扱う、生物学、農学、医科学の基礎から応用にわたる幅広い分野に大きなインパクトを与えるものです。
今後の期待
今回、ブフネラ以外の細菌から水平転移した遺伝子が、ブフネラの維持に利用されているという、予想を超える新知見を得ることができました。ではブフネラは、その進化過程において、ミトコンドリアや葉緑体のように、遺伝子を宿主のゲノムへ移すことはなかったのでしょうか?疑問はさらに広がります。現在、研究ユニットは、国内外の複数の研究チームと合同で、エンドウヒゲナガアブラムシ(図1)の全ゲノム配列の解析を展開しています。これにより、アブラムシのゲノム全体をくまなく調べることが可能になり、ブフネラからの遺伝子転移の有無については、近く決着すると考えています。
また、アブラムシは、重要な農業害虫種を多数含みますが、いずれもブフネラなしでは繁殖できません。共生系の存立基盤をより深く理解し、これを標的とすれば、生態系に負荷のかからない、安全な害虫防除法の開発につながると期待できます。
発表者
理化学研究所
宮城島独立主幹研究ユニット
ユニット研究員 中鉢 淳(なかばち あつし)
Tel : 048-467-9332 / Fax : 048-467-9329
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.相利共生細菌
複数種の生物が緊密な結びつきを保ちながら、同所的に生活する現象を共生と呼ぶが、これは大きく相利共生(双方の生物種が利益を得る)、片利共生(一方が利益を得るが、他方には影響がない)、寄生(一方が利益を得ると同時に他方が不利益を被る)の3つに分類される。すなわち相利共生細菌とは、共生関係によって、細菌と宿主の両者が利益を得るものを指す。ここではアブラムシに依存しながら、アブラムシに対して栄養分を供給してその繁殖を支える「ブフネラ」のこと。 - 2.体腔
動物の体壁と消化管との間の空所、空隙で、この中にさまざまな内臓を納める。動物の体制上、体腔内が「体内」であるのに対し、消化管内はあくまで「体外」である。 - 3.菌細胞
共生微生物を収納、維持するために分化した細胞で、さまざまな系統の昆虫など、無脊椎動物でしばしば見られる。 - 4.ブフネラ
Buchnera aphidicola (ブフネラ・アフィディコラ)。γ-プロテオバクテリア綱・腸内細菌目・腸内細菌科に属する細菌。これまでに4種のアブラムシに由来するブフネラの全ゲノム塩基配列が決定されており、そのゲノムサイズは420kb~650kbで、コードされている遺伝子は400~600程度。近縁な自由生活性の細菌である大腸菌のゲノムサイズが4Mbで、遺伝子数が4,000ほどであるのと比べて、著しくゲノムサイズが小さくなっているのが分かる。ちなみに生物界最小のゲノムは、キジラミの菌細胞内にすみ、やはりγ-プロテオバクテリア綱に属するカルソネラ( Carsonella ruddii; カルソネラ・ルディアイ)のもので、そのサイズは160kb (Nakabachi et al., Science, 2006; 2006年10月13日プレスリリース)。 - 5.垂直感染
微生物が、宿主から直接その子孫に伝播される感染様式。これに対して個体間で起こる、より一般的な感染は水平感染とよばれる。 - 6.水平転移
生物の遺伝子は、通常親から子へと伝えられていくが、これに対して親子関係にない生物体のゲノムの間で遺伝物質が移ること。(「水平感染」とはまったく別の概念であることに注意。)単細胞生物同士の間では比較的頻繁に起こることが知られているが、動物などの多細胞生物ではごくまれにしか見られない。これは、単細胞生物では、ゲノムが外来のDNA断片にさらされる頻度が高いばかりでなく、一旦そうした断片がゲノムに挿入されると、そのまま次世代に受け渡されるのに対し、多細胞生物では、細胞は体細胞と生殖細胞に分かれており、外来DNA断片が次世代に受け渡されるためには、体内に収納されたごく少数の生殖細胞の染色体に挿入されなければならないためである。さらに、細菌から多細胞生物に水平転移したDNA断片が「遺伝子」として働くには、 ※7に示すようにもうひとつの高い壁が存在する。 - 7.真核生物、原核生物
細胞核を持つ生物を真核生物、持たない生物を原核生物という。細菌が原核生物であるのに対し、動物を含む多細胞生物はすべて真核生物である。細胞の構造だけでなく、遺伝子構造、遺伝子発現機構なども両者の間で異なる。遺伝子発現機構が異なることから、細菌から水平転移したDNA断片が、真核生物内で遺伝子として発現、機能するためには、真核生物型の遺伝子構造を獲得する必要がある。この障壁があるため、細菌から真核生物に水平転移したDNA断片のうち、ほとんどは遺伝子として機能することなく、壊れていく一方であることが知られている。 - 8.オルガネラ化
ミトコンドリアや葉緑体といったオルガネラは、10億年以上前に真核生物の共通祖先に侵入、共生を開始した細菌に由来する、という「細胞内共生説」が現在広く受入れられている。これらのオルガネラは独自のゲノムを持つが、そのサイズは著しく縮小しており、大部分の遺伝子は宿主の核ゲノムに移行している。これより起源が新しく、1~2億年前に動物との共生を開始したのが、ブフネラやカルソネラといった菌細胞内共生細菌である。菌細胞内共生細菌はゲノムサイズが縮小し、宿主細胞への依存度が高いという点においてオルガネラに似るが、これまで遺伝子を宿主ゲノムに移行させているとの証拠は得られていない。 - 9.エクソン、イントロン、スプライソソーム型イントロン
遺伝子が発現する際、DNA上の遺伝情報はまずRNAへと転写されるが、転写された配列のうち一部はRNAの成熟の過程で除去される。遺伝子上で、この除かれる配列に対応する領域をイントロンとよび、成熟後もRNAとして残る配列に対応する領域をエクソンという。イントロンにはいくつかの型があるが、スプライソソームと呼ばれる構造により切り出されるものをスプライソソーム型イントロンといい、真核生物だけで見られる。 - 10.分子系統解析
あらゆる生物は遺伝子の本体である核酸と、それに基づくタンパク質を持つが、これらの塩基配列や、アミノ酸配列などを比較することで、生物間の系統関係や遺伝子の進化過程を推定する手法。 - 11.リアルタイム定量RT-PCR
試料中のRNAの存在量を定量する手法。試料から抽出したRNAを逆転写してcDNAとし、これを鋳型としたポリメラーゼ連鎖反応を行いながら、増幅産物をリアルタイムで追跡し、増幅率に基づいてcDNAの存在量を算出する。 - 12.ムレイン
ペプチドグリカンともいう。細菌の細胞壁の基本骨格をなす化合物で、多糖鎖に比較的短いペプチド鎖が結合した構造を持つ。 - 13.ボルバキア
Wolbachia。α-プロテオバクテリア綱・リケッチア目・アナプラズマ科に属する細菌。昆虫を含む節足動物や線虫に感染するが、宿主との関係は相利共生、片利共生、寄生とさまざまである。 - 14.リケッチア
Rickettsia。狭義のリケッチアはα-プロテオバクテリア綱・リケッチア目・リケッチア科・リケッチア属に属する細菌を指すが、アブラムシの lcdA遺伝子はアナプラズマ科のボルバキアやリケッチア科・オリエンチア属のツツガムシリケッチア( Orientia tsustsugamushi; オリエンチア・ツツガムシ)といった細菌の近縁種に由来すると考えられる。

図1 エンドウヒゲナガアブラムシ(撮影:中鉢淳)
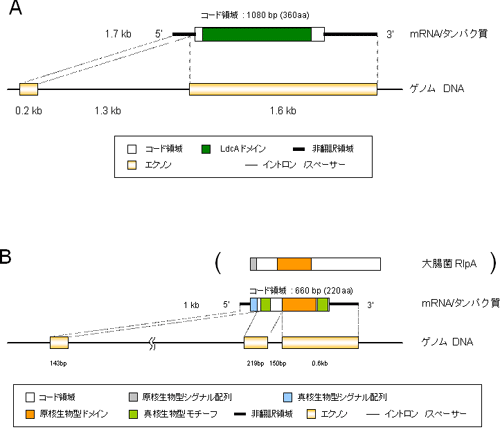
図2 水平転移遺伝子の構造
A:ldcA遺伝子 B:rlpA遺伝子
ldcA遺伝子、rlpA遺伝子ともに、スプライソソーム型のイントロンを持つ。またアブラムシのレアリポプロテインAは原核生物型の主要ドメインと真核生物型のモチーフから構成されるキメラ状の構造になっているのが分かる。括弧内は大腸菌のレアリポプロテインA。
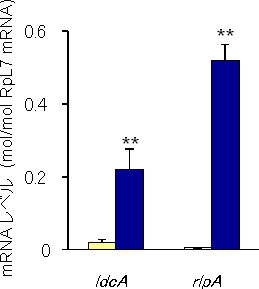
図3 水平転移遺伝子の転写産物レベル
白色カラム:体全体 青色カラム:菌細胞
水平転移遺伝子のmRNAコピー数を、リボソームタンパク質をコードするmRNAのコピー数で校正したもの。ldcA遺伝子、rlpA遺伝子ともに、菌細胞で著しく発現が亢進しているのが分かる。