2009年9月7日
独立行政法人 理化学研究所
遺伝子の働きを決めるヒストンアセチル化をリアルタイムで観察
―蛍光プローブ「Histac」を作製、Histac遺伝子導入で生きた細胞の様子を可視化ー
ポイント
- 生きた細胞内でヒストンH4の高アセチル化を検出する蛍光プローブを開発
- 細胞分裂時、ヒストンH4の脱アセチル化をリアルタイムで観察
- 各種の脱アセチル化酵素阻害剤の作用を可視化、がん治療薬の評価なども可能に
要旨
独立行政法人理化学研究所(野依良治理事長)は、染色体を構成するタンパク質の1つで、DNAが巻きついているヒストンがアセチル化される様子を、生きたままの細胞内でリアルタイムに観察することができる蛍光プローブ「Histac;ヒスタック」の開発に成功しました。理研基幹研究所吉田化学遺伝学研究室の佐々木和樹基礎科学特別研究員(現・脳科学総合研究センター客員研究員)、吉田稔主任研究員らと仏・国立保健医学研究所(INSERM;Institut National de la santé et de Ia Recherche Médicale)のサディ・コシュビン博士(Saadi Khochbin)との共同研究によるものです。
ヒストンのアセチル化やDNAのメチル化など、タンパク質やDNAの化学修飾は、遺伝子機能を変化・記憶し、その機能を子孫や娘細胞に伝えます。この現象はエピジェネティクスと呼ばれ、生命科学研究の大きなターゲットとなっています。ヒストンアセチル化はエピジェネティクスの調節を担う中心的な化学修飾で、エピジェネティクスによる細胞機能の記憶は、環境ストレスや老化などによって変化すると考えられ、その異常が、がんを含むさまざまな疾患の発症とかかわることが明らかになりつつあります。
一般的に、ヒストンの高アセチル化領域は遺伝子の転写活性化、低アセチル化領域は転写不活性化と相関することが知られ、ヒストンアセチル化は、ヒストンアセチル化転移酵素(HAT)とヒストン脱アセチル化酵素(HDAC)によってダイナミックに制御されています。しかし、これまでヒストンH4※1のアセチル化の様子を、生きた細胞内で検出することはできませんでした。
研究グループは、蛍光共鳴エネルギー移動(FRET)※2を利用してさまざまな試行錯誤を行い、蛍光プローブ「Histac」の作製に成功し、Histac遺伝子を導入した培養細胞を蛍光顕微鏡で観察して、生きた細胞内のヒストンH4のアセチル化の変化を観察することに成功しました。Histacを用いて細胞分裂の過程を観察したところ、細胞分裂の際に、一過的にヒストンH4が脱アセチル化されている様子を観察することができました。また、各種HDAC阻害剤を加えたところ、阻害剤ごとにアセチル化の反応速度に差があることが分かりました。
HDAC阻害剤はすでに抗がん剤として利用されており、Histacは今後、がん治療薬の評価、細胞の分化誘導やiPS細胞作製の際のアセチル化の動態の観察などに役立つと期待できます。
本研究成果は、米国科学アカデミー紀要『Proceedings of the National Academy of Sciences of the United States of America: PNAS』オンライン版に8月31日の週に掲載されました。
背景
高等生物のDNAは、ヒストンと呼ぶ球状のタンパク質複合体に1.65回転巻きつき、この複合体を基本単位として存在しています。ヒストンには、炭素2個、酸素1個、水素3個からなるアセチル(CH3CO)基が結合し、ヒストンアセチル化と呼ぶ化学修飾が起こります。ヒストンアセチル化は、ヒストンアセチル化転移酵素(HAT)とヒストン脱アセチル化酵素(HDAC)によって制御され、遺伝子発現のON/OFFのメインスイッチになっていると考えられています。近年、遺伝子発現調節機構の詳細が解明され、ヒストンアセチル化は、DNAのメチル化などほかの修飾とともに、DNAとは異なる遺伝暗号(エピジェネティックコード)として機能し、細胞が自分自身の素性を覚えて、分裂後も同様の機能を維持するための記憶装置として働いていることが明らかになってきました。このことは、組織が分化していく発生過程で、特定の遺伝子群の働きを決定し、正常な組織の機能を維持するために、ヒストンアセチル化が重要であることを示しています。一方、これを人為的に操作すると、胚の初期化やiPS細胞の作製にも有用であることが明らかになってきました。さらに、がんやメタボリックシンドロームなど、加齢、ストレス、生活習慣などに伴って起こる病気の多くにエピジェネティクスの異常が観察され、疾患の原因の1つであると考えられるようになってきました。そのため、治療薬としてヒストンアセチル化やDNAメチル化を調節する化合物が期待され、現在世界中で活発に創薬研究が行われています。すでに米国では、HDAC阻害薬が、抗がん剤として承認されるに至っています。
従来、ヒストンアセチル化を観察するための検出方法は、放射能標識するか、特異的な抗体を用いる方法に限られていました。これらの方法では、細胞を破砕して検出するか、いったん細胞を固定・染色するため、アセチル化/脱アセチル化を細胞が生きたままリアルタイムで観察することはできませんでした。そのため、細胞分裂、発生分化、細胞の老化や細胞死など、さまざまな局面で劇的に変化することが予想されるヒストンアセチル化の動態をとらえることは、非常に困難でした。
研究手法と結果
研究グループは、ヒストンアセチル化の動態の観察に、細胞内のカルシウムやタンパク質リン酸化の検出などに用いられている蛍光共鳴エネルギー移動(FRET)を利用できないかと考えました。この方法を用いるためには、アセチル化されたヒストンH4を特異的に認識して、強く結合するタンパク質が必要です。そこで仏・国立保健医学研究所のKhochbin博士が発見した、精巣で特異的に発現しているBRDTというタンパク質に着目しました。BRDTはアセチル化ヒストンに結合することが予想されるブロモドメイン※3という構造を2つ持ちます。実際、BRDTを培養細胞に発現させて、HDAC阻害剤であるトリコスタチンA(TSA)を加えてヒストンのアセチル化を上昇させると、BRDTが存在するときだけ染色体の凝縮が観察されることから、BRDTは高アセチル化ヒストンに強い結合能力があることが分かっていました。そこで、Khochbin博士と共同研究を行い、BRDTのブロモドメインがアセチル化ヒストンに特異的に結合するメカニズムを解析しました。その結果、コアヒストンと呼ぶ4種類(H2A、H2B、H3、H4)のヒストンのうち、ヒストンH4のアセチル化のみをBRDTは認識することが分かりました。ヒストンH4は、5番目、8番目、12番目、16番目に存在している4箇所のリジン(K5、K8、K12、K16)がアセチル化されることが知られていますが、さらに詳細に調べると、BRDTは4箇所のうち5番目と8番目のリジン(K5、K8)が両方ともアセチル化したときにだけ、ヒストンH4と結合するということが分かりました。そこで、蛍光タンパク質Venus、BRDTのブロモドメイン、20個のアミノ酸からなる柔軟なリンカー配列、ヒストンH4および蛍光タンパク質CFPを順番に直列でつなぎ、FRETの変化によってヒストンH4のアセチル化の変化を検出できるプローブを設計し、「Histac」と命名しました(図1)。
まず、このHistacが目的の通りに働くかどうかを調べるために、Histacの機能部位のアミノ酸をほかのアミノ酸に置換した変異体を作製して検証しました。その結果、ヒストンH4のK5がアセチル化されるためにはK12がアセチル化されていることが必要であることを発見しました。メカニズムの解明はこれからの課題になりますが、この結果は、K5、K8、K12、K16が、お互いの部位間でアセチル化の情報を伝達していることを示しており、エピジェネティクス研究に新しい展開をもたらす可能性が見えてきました。
次に、Histacを発現している細胞を蛍光顕微鏡下で培養し、HDAC阻害剤によりアセチル化を促進させてHistacの応答を観察したところ、ヒストンH4がアセチル化されるにつれて、核で発現しているHistacに含まれているCFPとVenusの蛍光強度の比(疑似カラーで表示)が変化することを確認できました。(図2)。
また、細胞分裂の際、一過的にヒストンH4が脱アセチル化された後、分裂を終えて間期になるにつれて、再びアセチル化されていく様子も観察しました。従来は、固定した細胞間での比較しかできなかったため、細胞分裂の過程でヒストンH4のアセチル化が変化するのか否か意見が分かれていましたが、今回、単一細胞で連続して観察することで、細胞分裂の際に脱アセチル化が起こっていると決着を付けることができました。
さらに、トリコスタチンA、CHAP31、SCOP304、SCOP402、FK228など各種HDAC阻害剤によって誘導されるヒストンH4の高アセチル化を観察したところ、各阻害剤によってアセチル化の反応速度に差があることも初めて分かりました。
今後の期待
HDAC阻害剤は抗腫瘍活性を持つことが広く知られており、臨床試験まで進んでいる化合物も多数あります。関連疾患はがんだけでなく、難病とされるアルツハイマー病、ハンチントン病などの神経変性疾患などにもHDACの関与が報告されているため、HDAC阻害剤はさまざまな治療薬への可能性を秘めていると考えています。今回開発したHistacは、生きた細胞内でHDACの阻害活性を評価できるため、治療薬の開発にも役立つと期待されます。また、細胞の分化や初期化の過程ではヒストンアセチル化の様子が変化すること、iPS細胞ではヒストン修飾などのエピジェネティクスの情報がリセットされていることなどが知られています。今回開発したプローブ「Histac」は、ヒストンアセチル化がこのような過程でどのように変化していくのかを明らかにすることができると考えています。
発表者
理化学研究所
基幹研究所 吉田化学遺伝学研究室
主任研究員 吉田 稔(よしだ みのる)
Tel: 048-467-9516 / Fax: 048-467-4676
脳科学総合研究センター
先端基盤技術研究コア細胞機能探索技術開発チーム
客員研究員 佐々木 和樹(ささき かずき)
Tel: 048-462-1111(代表) 内線:8002
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.ヒストンH4
真核生物の染色体は、主にヒストンと呼ばれるタンパク質とDNA によって構成されている。染色体は、ヒストンH2A、H2B、H3、H4のそれぞれ2分子からなる八量体に、DNAが左巻きに1.65回巻きついた複合体を基本単位として存在している。 - 2.蛍光共鳴エネルギー移動(FRET)
励起スペクトルと蛍光スペクトルに重なりを持つ2つの蛍光色素間の距離が近づくと、共鳴により励起エネルギーが直接移動し、他方の蛍光色素から蛍光が放出される現象。 - 3.ブロモドメイン
ヒストンアセチル化転移酵素(HAT)によってアセチル化を受けたリジン(塩基性アミノ酸の1種)に対して特異的に結合するタンパク質ドメイン。主にアセチル化されたヒストンを認識するHATや、転写因子などに含まれており、それらがブロモドメインを使ってアセチル化された場所に集合するために働くと考えられている。
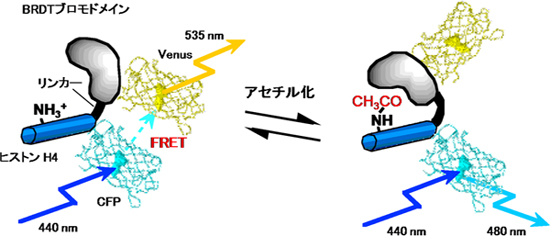
図1 Histacの原理図
左図:アセチル化前のHistacの状態。VenusとCFPが近い位置にあるため、黄色(535nm)の光を発する。
右図:アセチル化後のHistacの状態。VenusとCFPが遠い位置にあるため、シアン色(480nm)の光を発する。
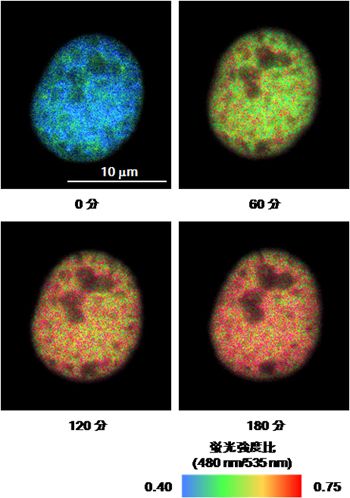
図2 HDAC阻害剤TSAを加えた後のHistacの応答
図はCFP(480nm)とVenus(535nm)の蛍光強度の比(480nm/535nm)を擬似カラー表示で示しています。アセチル化されるにつれて核の色が青から赤に変化していきます。