2010年3月18日
独立行政法人 理化学研究所
「植物の体内時計」に関与するタンパク質の生化学的機能を発見
-花芽の形成や組織サイズを制御する「植物の体内時計」形成メカニズムの理解へ-
ポイント
- 時計関連タンパク質「PRR9、PRR7、PRR5」の転写抑制機能を突き止める
- 「PRR」が作り出す16時間の期間が、植物の24時間周期形成の鍵に
- 体内時計の制御による、バイオマス増産やストレス耐性品種開発へ手がかり
要旨
独立行政法人理化学研究所(野依良治理事長)は、「植物の体内時計(概日時計)※1」に関与する3つのタンパク質のPRR※29、PRR7、PRR5が、転写※3活性を抑制する機能を持ち、特に、明け方に機能することで体内時計への関連が知られているCCA1遺伝子※4とLHY遺伝子※4の転写活性を抑制することを見いだしました。さらに、これら3つのタンパク質が発現する各々の時間の特定に成功し、16時間にわたる周期的な転写抑制を発揮していることを突き止めました。これは、理研植物科学研究センター(篠崎一雄センター長)生産機能研究グループ(榊原均グループディレクター)の中道範人基礎科学特別研究員、木羽隆敏研究員、国立大学法人名古屋大学農学部の水野猛教授、米国ロックフェラー大学のロザーナ・ヘンリクエス博士、ナムハイ・チュア教授との共同研究による成果です。
地球上の多くの生物は、体内時計を使うことで、地球の自転・公転に起因する昼夜や季節変動という環境変化に適応して生きています。近年の分子遺伝学の発達によって、植物の体内時計に関与する遺伝子が見つかってきましたが、実際に体内時計を作り出すメカニズムはいまだに謎です。
研究グループは、シロイヌナズナ※5のPRR9、PRR7、PRR5というタンパク質の機能解析やアミノ酸配列の解析を行い、これらには共通して転写を抑制する機能があることを突き止めました。さらに、生体内では、PRR9、PRR7、PRR5それぞれのタンパク質が、連続的にほかの時計関連遺伝子であるCCA1遺伝子とLHY遺伝子の転写調節にかかわる領域に結合し、転写活性を抑制していることを明らかにしました。この結合時間帯は、朝から夜中までの16時間にわたっており、この期間が体内時計の24時間の周期を作る上で重要だと考えられます。PRRそれぞれのタンパク質の生化学的な機能の情報を基に、PRRタンパク質の機能を「人工的にデザイン」する、または自然界から「変異型PRRを探索」することで、効率的な花芽形成やバイオマスの増産、ストレス耐性の付加を可能にすると期待できます。
本研究成果は、米国の科学雑誌『The Plant Cell』オンライン版に近く掲載されます。
背景
地球の自転・公転運動から生まれる昼夜や、季節変動という環境変化に対応するために、多くの生物は周囲を取り囲む環境の時間を予期するシステム「体内時計」を備えています(図1A)。この自転・公転のもと、太陽からの限られた光を有効に利用するために、植物は日の出を予期して、夜明け前から光合成の準備を始めます。また、繁栄に最も都合の良い季節に一斉に開花する現象も、体内時計を使った日長測定によって支えられています。さらに、近年の遺伝子探索の網羅的な研究手法により、体内時計は従来考えられてきた以上にさまざまな生理現象の「時間的な」制御を行っていることが分かってきました(図1B)。
この10年間で、モデル植物であるシロイヌナズナについては、その体内時計に関連するさまざまな遺伝子、例えばCCA1、LHY、TOC1(=PRR1)、PRR9、PRR7、PRR5などが同定され、体内時計の分子遺伝学的な研究が急速に進みました。その中でもPRR9遺伝子、PRR7遺伝子、PRR5遺伝子の重複的な欠損は、体内時計の作り出す「概日リズム」が消失するほどで、これらがコードするタンパク質PRR9、PRR7、PRR5は、それぞれ非常に重要な役割を持っていることが知られています。各タンパク質は、N末端※6にPRドメイン※7を、C末端※6にCCTモチーフ※8を持ち、細胞核内に局在していることは分かっていましたが、その生化学的機能については不明でした。そのほかの時計関連遺伝子がコードするタンパク質にも生化学的機能の分からないものが多く、体内時計の形成についてはいまだに謎が多い状態です。
研究手法と成果
(1)タンパク質PRR9、PRR7、PRR5の転写抑制機能の発見
研究グループは、PRR9、PRR7、PRR5の生化学的機能の情報を簡便に得るために、遺伝子銃法※9を用いて、それぞれのタンパク質を発現する3種類のベクターを、別々に植物体に導入し、CCA1遺伝子、LHY遺伝子、TOC1遺伝子の転写活性を調べました。その結果、明け方に転写されるCCA1遺伝子とLHY遺伝子の転写活性を抑制することを見いだしました。また、シクロヘキシミドという薬剤を用いて、生体内でのすべてのタンパク質の新規合成を阻害させましたが、この条件でもPRR5タンパク質は、CCA1遺伝子とLHY遺伝子の転写活性を抑制することが分かりました。つまりPRR5タンパク質によるCCA1遺伝子とLHY遺伝子の転写抑制は、直接的(新規なタンパク質の合成を必要としない)であると分かりました。さらに、PRR9、PRR7、PRR5タンパク質のアミノ酸配列を解析したところ、3種のPRRタンパク質に共通の新規な転写抑制モチーフを発見しました(図2)。これらの一連の解析より、PRR9、PRR7、PRR5タンパク質には転写活性を抑制する機能があることを突き止めました。
(2)PRR9、PRR7、PRR5の働く時間帯の特定
これまで、PRR9、PRR7、PRR5のそれぞれのタンパク質は、DNAに結合するとは考えられていませんでした。しかし、クロマチン免疫沈降(Chromatin Immunoprecipitation; ChIP)※10を用いて解析すると、生体内でこれらのPRRタンパク質群が、CCA1遺伝子とLHY遺伝子の転写調節にかかわるDNA領域に結合していることを見いだしました。また、この結合している時間帯を調べるために、24時間にわたる経時的なChIP解析を行ったところ、PRR9は朝、PRR7は朝から夜中、PRR5は夕方から夜中に結合しており、これらの時間帯は、CCA1遺伝子とLHY遺伝子の転写活性が抑制されている時間帯と一致していることが分かりました(図3A、B)。さらにPRR7遺伝子とPRR5遺伝子の両方を欠損すると、CCA1遺伝子とLHY遺伝子の「夜間」の転写抑制が、PRR9遺伝子とPRR7遺伝子の両方を欠損すると「昼間」の抑制がそれぞれ解除されました(図3C、D)。つまり、CCA1遺伝子とLHY遺伝子の16時間ほどにわたる長時間の転写抑制状態には、PRR9、PRR7、PRR5のタンパク質が、それぞれ順番に転写調節領域に結合することが重要であると分かりました。
体内時計の周期長である「約24時間」は、PRR遺伝子の欠損によって大きく変わることからも、今回発見した16時間というPRRタンパク質が機能する期間は、「約24時間」周期の形成にとって重要な要素であることが考えられます。
今後の期待
近年の遺伝子探索の網羅的な研究手法により、植物の体内時計は、従来考えられてきた以上にさまざまな生理現象の時間的な制御を行っていることが分かってきました。シロイヌナズナのPRR9、PRR7、PRR5の各遺伝子は、概日リズムを調節することで、花芽の形成、組織サイズ(バイオマス)の制御、ストレス応答にかかわる遺伝子群の発現制御、ミトコンドリアや葉緑体といった細胞内器官にある代謝物経路のホメオスタシス※11に関与することが分かっています(2009年4月7日プレス発表)。今回発見したPRRタンパク質の生化学的な機能の情報を基に、PRRタンパク質の機能を「人工的にデザイン」する、または自然界から「変異型PRRを探索」することで、作物をはじめとする有用植物の効率的な花芽形成、バイオマス増産、ストレス耐性の付加を可能にすると期待されます。
発表者
理化学研究所
植物科学研究センター 生産機能研究グループ
グループディレクター 榊原 均(さかきばら ひとし)
植物科学研究センター 生産制御研究チーム
基礎科学特別研究員 中道 範人(なかみち のりひと)
研究員 木羽 隆敏(きば たかとし)
Tel: 045-503-9576 / Fax: 045-503-9609
お問い合わせ先
横浜研究推進部Tel: 045-503-9117 / Fax: 045-503-9113
報道担当
理化学研究所 広報室 報道担当Tel: 048-467-9272 / Fax: 048-462-4715
補足説明
- 1.体内時計(概日時計)
多くの生物で見られる遺伝的に組み込まれた時計。特に周期が約24時間のものを概日時計という。概日時計が生み出すリズムは概日リズムといい、時間の手がかりとなる環境変化(光環境や温度)が一定であっても、自律的な周期が観察できる。 - 2.PRR
PSEUDO-RESPONSE REGULATORの略称。微生物や植物に普遍的な情報伝達機構として知られるタンパク質リン酸リレー系(His-Aspリン酸リレー系)の出力タンパク質「RR(レスポンスレギュレータ)」の疑似体。
PRRはRRが持つリン酸受容能力が無くなっているので、疑似(pseudo)レスポンスレギュレータと呼ばれる(出典:蛋白核酸酵素vol.47 No.11 水野猛著、(2002))。PRRは植物に普遍的に存在する。シロイヌナズナでは、PRR9、PRR7、PRR5、PRR3、TIMING OF CAB EXPRESSION 1(TOC1=PRR1)が体内時計に関与している。 - 3.転写
多くの遺伝子の機能は、DNAからのmRNAへの「転写」、mRNAからタンパク質への「翻訳」を経て発現する。転写のステップは、積極的に制御を受けている例が多いが、それを負に制御することを転写抑制という。 - 4.CCA1遺伝子、LHY遺伝子
それぞれ CIRCADIAN CLOCK-ASSOCIATED 1 と LATE ELONGATED HYPOCOTYL の略称。どちらも相同性の高いMYB型転写因子をコードしている。明け方に発現してTOC1遺伝子の転写を抑制するなどを行い、体内時計の形成にかかわる。両遺伝子の機能喪失は概日リズムの短周期化を引き起こす。 - 5.シロイヌナズナ
学名は Arabidopsis thaliana (L.) Heynh. アブラナ科シロイヌナズナ属の1年草で実験モデル植物であり、2000年にゲノム解読が行われた。全遺伝子数は約30,000個。 - 6.N末端、C末端
タンパク質はアミノ酸のポリマーであり、個々のアミノ酸は、アミノ基とカルボキシル基でペプチド結合をしている。このポリマーの末端の非結合型のアミノ基側をN末端、カルボキシル基側をC末端と呼ぶ。 - 7.PRドメイン
pseudo-receiverドメインの略。PRRに共通してみられるドメイン。PRR5とTOC1(PRR1)のPRドメインは、その選択的分解を担うZEITLUPEタンパク質と結合する。 - 8.CCTモチーフ
花芽の形成に重要なCONSTANS、いまだに機能未知のCONSTANS-LIKE 1(COL1)、体内時計に関連するTOC1のすべてのタンパク質に共通してみられるモチーフ。それぞれのタンパク質の頭文字をとって、CCTと呼ばれる。奇数番号のPRRにも共通してみられ、タンパク質間の結合に関与していると考えられている。 - 9.遺伝子銃法
導入したいDNAを金粒子にコーティングし、遺伝子銃(パーティクルガン)の物理的圧力によってその金粒子を生体に導入する方法。DNAにコードされた遺伝子の転写活性、機能などを簡便に観察することができる。 - 10.クロマチン免疫沈降(Chromatin Immunoprecipitation; ChIP)
ホルムアルデヒドでタンパク質とDNAを架橋し、抗体で目的タンパク質を免疫沈降させ、その沈降画分に含まれるDNAを検出する方法。転写因子とターゲット配列DNAの「生体内」での結合を見るために使う。 - 11.ホメオスタシス
生物の持つ恒常性の維持に関する性質のこと。生物を取り巻く外部環境と密接にやりとりをしつつ、生体内の状態が一定に保たれる性質を指す。例えば、体温や血圧、血糖値などが一定に保たれる(あるいは病気が治る)などもホメオスタシスと言える。米国の生理学者Walter B. Cannon(1871-1945)がこの概念を考え出した。
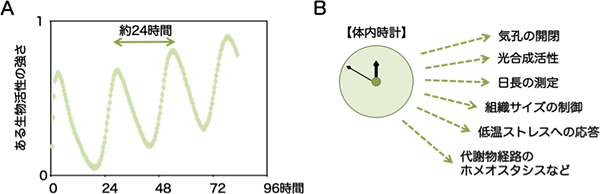
図1 体内時計(概日時計)のつくる概日リズム
(A)体内時計の作る概日リズムは、光や温度が一定である条件(環境の時間情報がない条件)でも約24時間の周期を示す。(出典:「マメから生まれた生物時計」田澤仁著、学会出版センター)
(B)植物の体内時計の制御する生理現象。上記以外にも、多くの現象が体内時計に制御されていることが知られる。
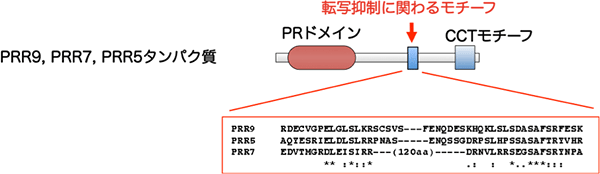
図2 PRR9, PRR7, PRR5タンパク質の機能発見
PRR9、PRR7、PRR5のそれぞれのタンパク質で発見した転写抑制にかかわるモチーフ(赤字)。モチーフのアミノ酸配列を下に表した。このモチーフは、多くの植物由来PRRタンパク質で保存されている。
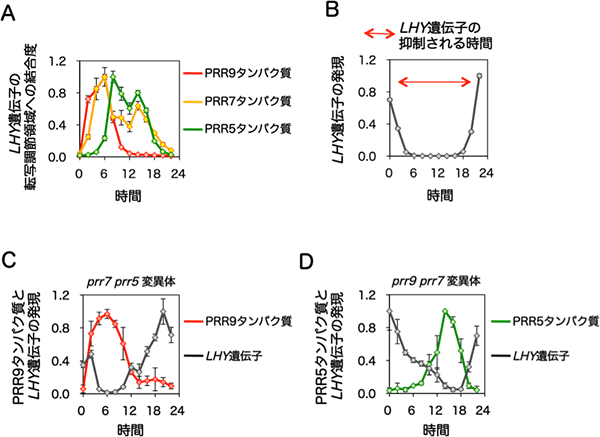
図3 PRR9、PRR7、PRR5タンパク質の機能する時間(日の出からの時間)の特定
(A)PRR9、PRR7、PRR5のそれぞれのタンパク質は、生体内でCCA1遺伝子とLHY遺伝子の転写調節領域に結合する。PRR9は日の出より2~8時間後に、PRR7は4~14時間後に、PRR5は8~16時間後に、という順番で結合する。図ではLHY遺伝子の転写調節領域への結合のみを示す。
(B)LHY遺伝子の発現パターン。PRR9、PRR7、PRR5のタンパク質がLHY遺伝子の転写調節領域に結合している時間(日の出より2~18時間後:赤矢印)ではLHY遺伝子の転写抑制が認められる。
(C)PRR7遺伝子とPRR5遺伝子の二重変異体(prr7 prr5)では、PRR9タンパク質のみが発現している(赤線)。この図ではLHY遺伝子(黒線)の転写の「夜間(日の出から12~18時間後)」の抑制が解除する。
(D)PRR9遺伝子とPRR7遺伝子の二重変異体(prr9 prr7)では、PRR5タンパク質のみが発現している(緑線)。この図ではLHY遺伝子(黒線)の転写の「昼間(日の出から4~12時間後)」の抑制が解除する。